|
|
Active Site of beta-Amylase |
 |
|||||||||||||
 |
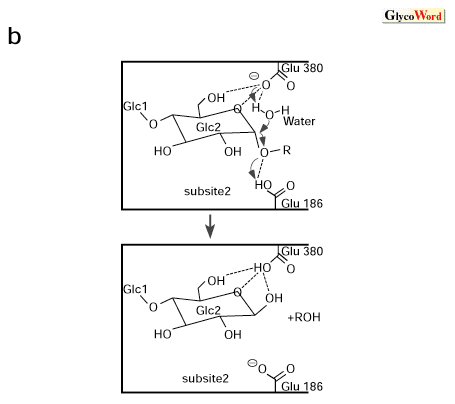
beta-Amylase catalyzes the liberation of maltose from the nonreducing ends of starch. The enzyme has a strict specificity to produce beta-anomeric maltose and has been classified as a typical inverting enzyme together with glucoamylase by Koshland. The enzyme has a molecular weight of 50-60 kDa and is distributed in higher plants such as soybean, sweet potato and barley, and in some microorganisms. The enzyme properties of bacterial beta-amylases are different from that of the plant enzyme in optimum pH and in their ability to digest raw starch granules. Since the three-dimensional structure of soybean beta-amylase was first determined, the structures of the enzymes from sweet potato, barley and Bacillus cereus have been clarified. It was found that Bacillus cereus beta-amylase contains a C-terminal starch binding domain instead of a long C-terminal loop found in the higher plant enzymes. The core structure of these beta-amylases are composed of (beta/alpha)8-barrel which has no similarity with that of the alpha-amylase family enzyme. The active cleft of beta-amylase is formed in the C-terminal side of the beta-barrel. Three long loops of L3, L4 and L5 surround the cleft and makes it a deep active site pocket. The catalytic mechanism of beta-amylase has been deduced from the structure of soybean beta-amylase complexed with maltose. Two maltose molecules bind in tandem on both sides of the cleavage point between subsites 2 and 3. The product from beta-maltose binds at subsite 1-2, so that one more maltose can bind in tandem at subsite 3-4. Figure 1-a shows the amino acid residues interacting with maltose. The glucose residues at subsites1-3 forms 8-10 hydrogen bonds with the protein residues. Although the glucose residue at subsite 4 makes only one hydrogen bond with His300, it has many van der Waals contacts with the surrounding hydrophobic residues. The flexible loop (residues 96-103) takes an open conformation in the apo enzyme and a closed conformation in the complex with maltose to interact with substrate. The most remarkable feature of the bound maltose conformation is that the torsion angle of the sugar rings are rotated almost 180o between subsites 2 and 3. Two side chains of Glu186 and Glu380 are involved in the catalytic process. Glu186 exists near the glycosidic bond to be cleaved, whereas Glu380 is located near the beta-anomeric O1 of product maltose at subsite 2. Figure 1-b shows the deduced catalytic mechanism of beta-amylase. Glu186 acts as a general acid and donates hydrogen to the glycosidic bond to be cleaved. On the other side, Glu380 acts as a general base and activates a water molecule, which attacks the C1 atom of a possible carbonium cation intermediate. The attack of the water molecule only from the Glu380 side gives strict product specificity for beta-anomeric maltose. The product maltose is released when the flexible loop opens. |
|||||||||||||
|
||||||||||||||
| Bunzo Mikami (Research Institute for Food Science, Kyoto University) |
||||||||||||||
|
||||||||||||||
| Jun. 15, 2000 | ||||||||||||||
|
|
||||||||||||||
|
||||||||||||||