

|
|
Structure and Catalytic Mechanism of alpha-Amylase Family Enzymes |
 |
|||||||||||||
 |
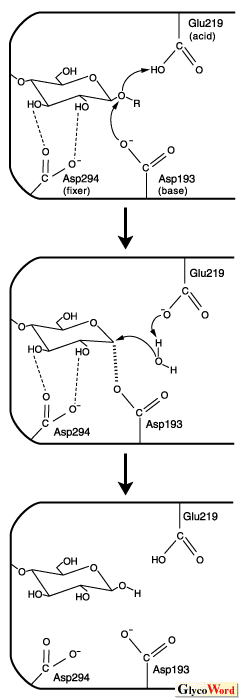
The enzymes whose structure has been determined by X-ray crystallography consistently possess a (beta/alpha)8 barrel supersecondary structure. The active site is always located at the C-terminal end of the 3rd and 4th beta-strands of the central beta-barrel. For these reasons, the catalytic mechanism of the hydrolysis of starch in the family enzymes can be considered as essentially the same. Furthermore in the three-dimensional structure, there exist several sub-domains in addition to the main domain, which are characteristic in the polypeptide chain foldings and relative positions with respect to the main domain. The role of the sub-domains is not yet fully understood. However, for enzymes capable of raw-starch binding, the sub-domain at the C-terminal end has been found to be responsible for this activity. The differences in the substrate specificities are related to the structures of the sub-domains that flank the main domain, forming altogether the active cleft. The roles of the three catalytic residues are discussed here according to the results obtained by the author’s group on the Pseudomonas stutzeri maltotetraose-forming alpha-amylase (G4-amylase), in which the complexes between mutant enzymes and the substrate have been analyzed (1,2). These mechanisms can be generalized in the alpha-amylase family enzymes. 1. Glu219 (Glu230: Taka-amylase) This residue is always conserved in the family enzymes (conserved region 3). However, there are no other residues conserved near this residue, making it the sole conservative. The carboxylate of this residue is believed to have a high pKa value caused by the environment, making it possible to easily liberate a proton on binding the substrate, followed by its addition to the glucosidic bond leading to cleavage (acid catalyst). 2. Asp193 (Asp206: Taka-amylase) X-ray structure analyses revealed that one carboxylic oxygen of this residue resides very close to the atom C-1 of the cleaving end glucose residue (position -1), suggesting that this residue is involved in the intermediate formation of the catalytic reaction (base catalyst/nucleophile). It is still under dispute whether the true intermediate takes a carbonium ion or undergoes covalent bonding with the carboxyl oxygen of this residue. Recently a paper has been published which describes the structure of a covalent adduct between sugar and the caboxyl oxygen (3). However, it may not be valid proof of what was the true intermediate. The author of this article believes that the true intermediate may rather take a somewhat intermediate form between two possible forms or take both forms cousecutively in the reaction time coordinates. 3. Asp294 (Asp297: Taka-amylase) Recently the author’s group has clarified the role of this residue by detailed X-ray structure analyses of the complexes between maltotetraose and five mutants of G4-amylase. These studies showed that the ring distortion of the glucose residue at position -1 which is consistently observed when this residue is aspartic acid, disappeared when it was altered to asparagine, adopting an approximate regular chair conformation. Furthermore these studies put forward evidences that the maltotetraose binds weakly in the case of asparagine. From these results, we can conclude that the role of this residue is to strengthen the substrate binding, giving rise to distortion of the sugar ring leading to the easy cleavage of the glucoside bond (fixer for catalysis) (2). Taken together the above discussions, a schematic picture showing a catalytic mechanism involving three residues is shown here. |
|||||||||||||
 |
||||||||||||||
| Yoshiki Matsuura (Institute for Protein Research, Osaka University) | ||||||||||||||
|
||||||||||||||
| Dec. 15, 1999 | ||||||||||||||
|
|
||||||||||||||
|
||||||||||||||

