|
|
グリコシラーゼの触媒機構 |
 |
||||||||||
 |
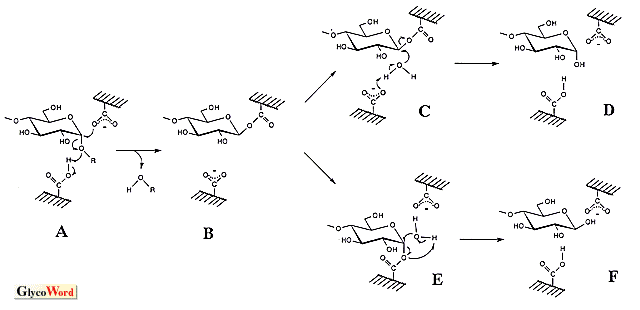
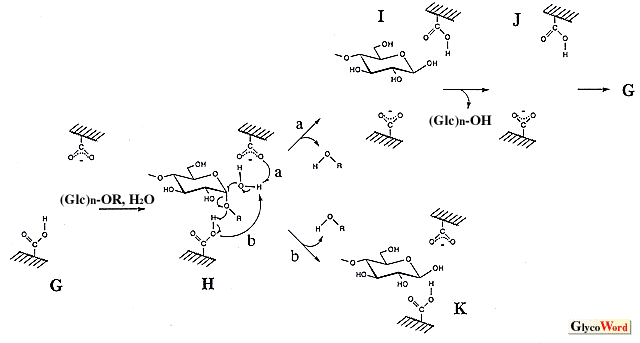
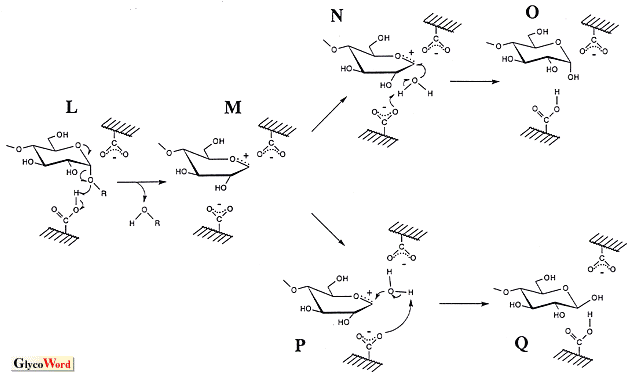
グリコシラーゼは、オリゴ糖、多糖や配糖体のグリコシド結合を加水分解する酵素の総称である。本稿ではシリーズの内容を考慮し、グルコース残基のアノメリック炭素とグルコシル結合の酸素間を切断するアミラーゼを中心に話を進める。 これらの酵素群(他のグリコシラーゼを含め)は、加水分解反応の他に糖転移反応や縮合反応をも触媒する。後の二反応は重合度の大きいオリゴマーを生成するため、良好な生理機能を示すオリゴ糖の合成に利用され、高い応用性を有する。糖転移作用は、グリコシル基をアルキル(或いは、アリル)水酸基へ転移する反応であり、反応機構は加水分解反応と同じであると考えられる(加水分解作用は水分子への転移反応とも捉えられる)。従って、加水分解反応の基質(供与体)が必須であり、受容体を選択することにより様々なオリゴ糖が得られる。高収率を望むならば、受容体濃度を高める必要がある(加水分解と競合するため)。一方、縮合作用は加水分解反応の逆方向に進行する脱水反応であり、加水分解反応の生成物が出発物質(基質)となる。糖転移作用によって形成されるグリコシル結合のアノマー構造は加水分解反応の生成物の立体配置と等しくなり、縮合作用では加水分解反応の基質の構造となる。 アミラーゼの加水分解反応の触媒機構については統一的な見解が得られておらず、二つの機構(オキソカルベニウムカチオン中間体を経由するSN1型反応と求核置換によるSN2型反応)が有力であり、両者を支持するデータが得られている。一方、加水分解作用における基質と生成物のアノマー構造の関係をみると、α-アミラーゼやα-グルコシダーゼなどの保持型酵素とβ-アミラーゼやグルコアミラーゼなどの反転型酵素がある。この保持・反転を説明するため、両反応機構から図に示したスキームが考えられている(アミラーゼはいずれもカルボキシル基を直接の触媒基としているようである。α-グルコシダーゼではイミダゾール基が候補になったことがあるが、プロトン化したカルボキシル基である)。 SN2型触媒反応ではアノマー構造の保持・反転機構を別個に考えねばならないようである。保持型酵素の場合、-COOHのグルコシル結合切断とともに、-COO- がアノメリック炭素を攻撃する(図のA)。共有結合を介したグルコシル酵素(B)を生じた後、水分子による2回目の反応(C)によりアノマー構造が保持される(D)。ここでSN2で保持・反転を統一的に理解する立場から反転型反応をみると、水分子の攻撃前に一方の-COO- がアノメリック炭素と一度反応する機構(E)となる。これは合計3回の求核置換を伴うことになり、不自然なスキームとなる。そこで、求核攻撃を1回で完了させる別の機構(H)が考えられている。一方、SN1型触媒反応では、-COOHによりグルコシル結合が切断された(L)後、生成されるオキソカルベニウムカチオン中間体はマイナスチャージしたカルボキシル基により安定化され(M、イオン的結合の形成)、これへのOH-付加方向によりアノマー保持・反転の仕組みが統一的に説明できる(それぞれ、NおよびP)。反転型酵素のスキームで、反応終了時にIの構造をみかけるが、これは水分子の活性化を-COO- が行う考えに基づく(Hの矢印aを経由する反応)。この場合、GとJの比較で分かるように反応の前後でカルボキシル基のプロトン化状態が逆転し、次の基質を受け入れる前にプロトン移動(J→G)が必要となる(Hの矢印bを経るならば、Kとなり、Gの構造が復元する)。SN2の3回求核置換型スキームやSN1型機構であっても、水分子のプロトン引抜を-COO- (EおよびP、Pは図の上側の-COO- )が行うのであれば、同様なプロトン化状態の逆転が生じ、プロトン移動が必要となる。これに関し、触媒基のpKe値によって溶媒中の水分子との間で移動がなされるとする説明もある。 アノマー構造保持機構を両スキームで比較すると、反応中間体(SN1ではオキソカルベニウムカチオン、SN2ではグルコシル基)とカルボキシル基との間の結合に大きな相違があることがわかる。すなわち、SN1型はイオン結合、SN2型は共有結合であり、その結合距離が重要となる。結合距離が一方に完全に片寄っているといずれかの反応機構を考えねばならなくなるが、そのような完全な片寄については疑問視する考えもある。すなわち、結合距離が両者の中間値であると、イオン的・共有的性質を併せ持つ結合となり(むしろ、化学結合を理解する上で自然な考え)、アノマー保持型アミラーゼの触媒機構を統一的に捉えることができるように思われる。反応素過程に生じる結合距離変化も想定でき、SN1型とSN2型の両機構を支持する実験データの存在もこの考えで説明できる。一方、反転機構についてはSN1とSN2は異なっている。Hにあるように活性化された水分子がアノメリック炭素に起こす求核攻撃と-COOHのグルコシド結合の酸素原子への反応が協奏的に進行し、加水分解反応が一挙に完了する。これはMのような中間体の存在を許さない。 最近、Kanekoらがドッキングモデルを用いて行った反応機構のシュミレーションに関する総説(J. Appl. Glycosci. 46, 187-197, 1999)も参考にされたい(極く短時間に生ずる酵素反応の素過程を実験的に直接捕捉することは困難であり、それを打開する有力な手段-シュミレーション-を用いて興味深い知見を解説している)。 |
||||||||||
|
|||||||||||
| 木村淳夫 (北海道大学・大学院農学研究科) | |||||||||||
|
|||||||||||
| 1999年 9月 15日 | |||||||||||
|
|
|||||||||||
|
|||||||||||