|
|
Receptors for the Oligosaccharide Signals |
 |
||||||||||||||||
 |
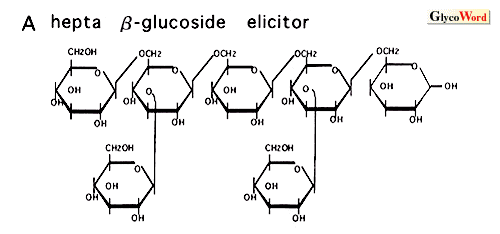
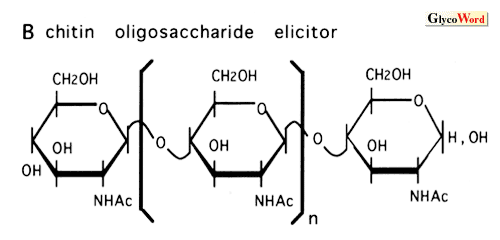
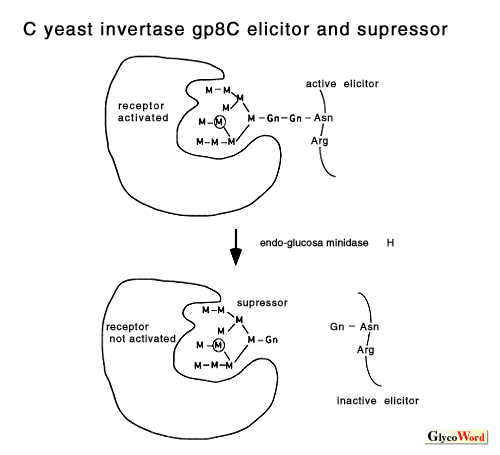
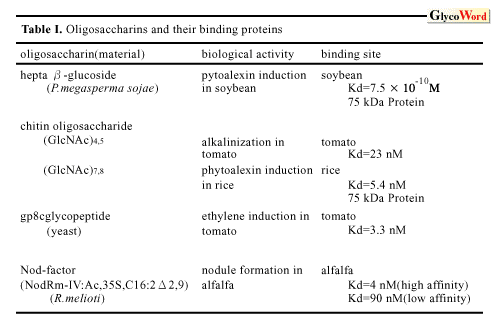
How do plant cells perceive and transduce the oligosaccharide signals? The highly specific structures and low concentrations required for the induction of plant defense responses by elicitor molecules strongly suggest the presence of a receptor molecule in the corresponding plants. Indeed, several high-affinity binding sites (putative receptors) for these elicitors have been identified in various plants by binding assay and affinity labeling. The best characterized elicitor binding protein is hepta beta-glucoside binding protein identified in the plasma membrane of soybean root cells. The protein, molecular weight 75 k, was purified to homogeneity and the cDNA was cloned and sequenced. The affinity of the binding site (Kd= 0.8 to 3 nM) for the elicitor was comparable to the concentration required for phytoalexin accumulation. Competitive inhibition by a group of chemically synthesized structural homologues of hepta beta-glucoside demonstrated a direct corelation between the binding affinity and the elicitor activity of the oligoglucosides tested. The results suggested that the binding site is a functional receptor for the elicitor signal. However, the predicted sequence of the binding protein had no homology with any known proteins and further characterization is necessary to determine its function in the signal transduction cascade. The binding sites for the chitin oligosaccharide elicitor have been studied in two plant systems, tomato and rice. In the rice system, the specificity and the sensitivity of the binding activity of the binding protein to the chitin oligosaccharides were in good agreement with the eliciting activity, suggesting that the protein is a functional receptor for the elicitor. It is probable that the 75 kDa hepta beta-glucoside binding protein and the similar sized chitin oligosaccharide binding protein of rice cells, which was identified by affinity labeling, belong to a family of receptors for carbohydrate elicitors. Cloning and characterization of the cDNA for this protein is now under way. The eliciting activity of chitosan requires relatively high concentrations and might be the result of disturbances of the membrane structure rather than a specific signal perception process. The specific binding site for gp8C elicitor, a glycopeptide of yeast invertase, was identified in tomato cell plasma membrane. Inhibition assay demonstrated that the gp8C and its glycan (suppressor of the eicitor) act as agonist and antagonist for the induction of the stress response by competing at this binding site. Recently, the binding protein was partially purified. The induction of the resistance of higher plants against pathogens may be highly dependent on the elicitor-receptor interactions. It is demonstrated that the binding proteins for the carbohydrate elicitors are present in many plant species, indicating that these binding proteins may be involved in the broad resistance rather than race-cultivar specific resistance. Studies on the Nod factor perception in legume plants have just started. The presence of a high-affinity binding site for the Nod factor of Rhizobium meliloti was demonstrated in the membrane enriched fraction of alfalfa, the only one high-affinity binding protein for Nod-factors reported so far. The results showed that the substitutions on the nonreducing sugar of the lipochitooligosaccharides are determinants for the high affinity binding to the binding protein. Further investigation is necessary to understand the function of the protein in the signal transduction cascade and in the host specificity of the Nod-factor. Oligogalacturonides elicit a variety of responses from defense against pathogen to morphogenesis in variouse plant systems. The structural requirements and the concentrations necessary for the activity are different in each case. In some cases, oligogalacturonides act antagonistic to auxin. In the tobacco thin cell layer explant system, the oligogalacturonides control organogenesis at concentrations as low as nanomolar levels. Characterization of oligogalacturonide receptors in this system should facilitate the elucidation of the mechanisms by which oligogalacturonides can show so many biological effects. It is not known how nanomolar concentrations of xyloglucan oligosaccharides can serve as a biological signal in plants. In the pea stem elongation system, the requirement for a specific fucosyl residue as well as the effectiveness at nM concentrations suggest the involvement of a specific receptor. However, the binding protein has not yet been reported. Studies on the signal transduction cascade downstream of the receptor, which results in the activation of the elicitor-responsive genes, are described in the following section. |
||||||||||||||||
|
|||||||||||||||||
| Figure 1. Microbial elicitors of plant defense responses | |||||||||||||||||
 |
|||||||||||||||||
| Yuki Ito (Department of Biotechnology, National Institute of Agrobiologycal Resources) | |||||||||||||||||
|
|||||||||||||||||
| Mar. 15, 2000 | |||||||||||||||||
|
|
|||||||||||||||||
|
|||||||||||||||||