|
|
ペプチド:N -グリカナーゼ(PNGase)と細胞質の遊離N 型糖鎖 |
|||||||||||||||||||||||
|
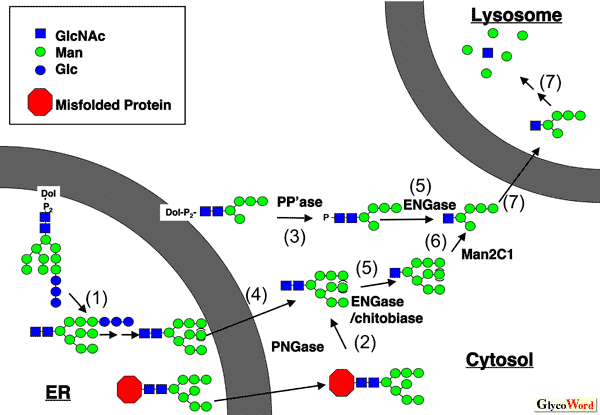
細胞質ペプチド:N -グリカナーゼ ペプチド:N -グリカナーゼ(PNGase)は糖タンパク質あるいは糖ペプチドからN 型糖鎖を遊離させる酵素であり、細菌あるいは植物由来の本酵素は糖タンパク質糖鎖の構造および機能研究の際の有用な試薬として用いられてきている。 一方で真核細胞の細胞質に広く存在するPNGaseは、現在ERにおいて新たに合成された異常糖タンパク質の品質管理機構の一つである小胞体関連分解(ERAD)に関わる分子であることが広く認知されるに至っている。この酵素は構造的に“トランスグルタミナ ーゼスーパーファミリー”に属する酵素で、システイン、ヒスチジンとアスパラギン酸からなる活性中心を持つ。哺乳動物のPNGaseオルソログはそのN末にPUBドメインと呼ばれる構造を持ち、様々なタンパク質間相互作用に関わっていることが予想されている。 一方、C端側のドメインは最近糖鎖結合部位であることが明らかとなった。これらのドメインは酵素のタンパク質複合体形成、および基質認識に重要な役割を果たしていることが示されつつある。本酵素の構造と生理機能についての詳細については、他稿を参照されたい(1-3)。 |
|||||||||||||||||||||||
|
遊離N 型糖鎖:生成と分解 PNGaseの細胞質における存在とERADへの関与は明らかとなったものの、そこで新たな疑問が生じる: PNGaseの反応によって遊離された糖鎖は一体どのような運命を辿るのだろうか?不思議なことに、新生 タンパク質の品質管理機構の一つである小胞体関連分解(ERAD)経路の発見以来、タンパク質の分解に関 しては研究が進んでいるが、一方で外された糖鎖の分解経路とその生物学的重要性についてはほとんど研究 の進展がないのが現状である。哺乳動物細胞においては、これまでエンド-β-N -アセチルグルコサミニダー ゼ(ENGase)とα-マンノシダーゼ(Mac2C1)という2つの細胞質グリコシダーゼが遊離糖鎖の分解に関わっ ていることが明らかにされている(図1)。この非リソソーム系の糖鎖の代謝は恐らく真核生物において最も 基本的な生物学的素過程であるにも関わらず未同定な酵素、トランスポーターが数多く存在することは注目に 値する(図1)。遊離糖鎖の生成、および代謝機構についてのより詳細は、我々の最近の総説を参考いただきたい(4)。 |
|||||||||||||||||||||||
|
|||||||||||||||||||||||
| 鈴木 匡 (大阪大学大学院 医学系研究科 生化学教室(21世紀COEプログラム)CREST/JST) | |||||||||||||||||||||||
|
|||||||||||||||||||||||
| 2007年4月19日 | |||||||||||||||||||||||
|
|
|||||||||||||||||||||||
|
|||||||||||||||||||||||