|
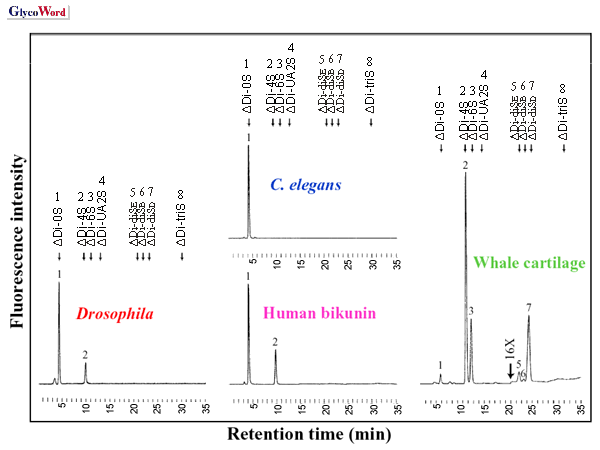
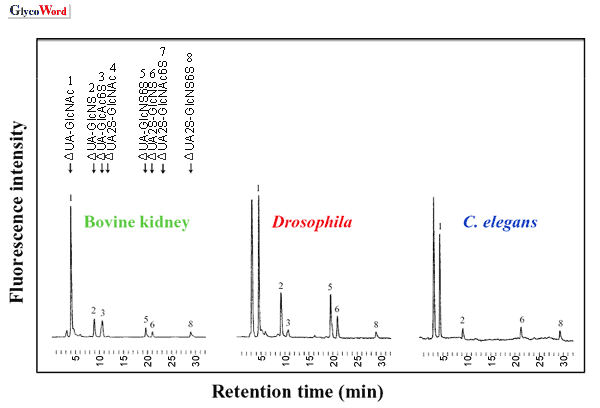
ヘパリナーゼによるショウジョウバエおよび線虫グリコサミノグリカンの消化によって,脊椎動物と同様の,N-, 2-O-,
6-O- 位が硫酸化された,non-, mono-, di-, tri- 硫酸化の不飽和二糖が生成した(Fig.
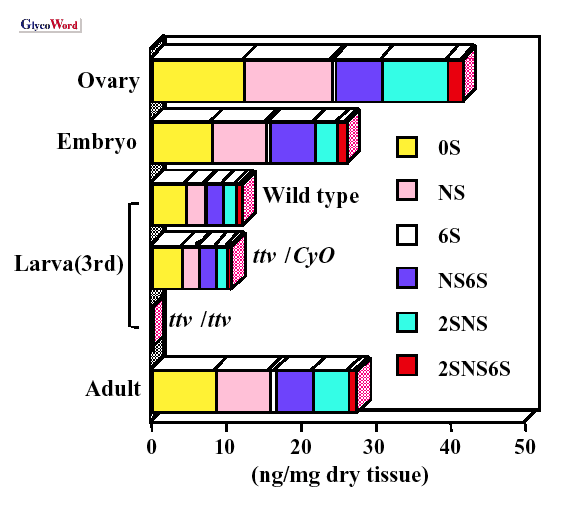
2)。グリコサミノグリカンの際立った特徴の一つは,組織の相違に応じて見られる構造多様性である。そこでショウジョウバエがヘパラン硫酸の構造と機能を研究するモデルシステムとなり得るかどうかを調べるために,われわれは,異なる発生段階と組織のヘパラン硫酸を分析した(2)。実際に,ショウジョウバエは組織および発生段階特異的なヘパラン硫酸の変化を示した(Fig.
3)。例えばΔUA-GlcNS6Sの割合は,幼虫や成虫に比べて胚で相対的に高くなっている。グルコサミンの6-O-硫酸は,発生を通じて成長因子のシグナル伝達を制御するため非常に重要である(3)。 |
|
|
| References |
(1) |
Selleck, S.B. (2000) Trends Genet. 16,
206-212
|
|
(2) |
Toyoda, H., Kinoshita-Toyoda, A., and Selleck, S.B. (2000)
J. Biol. Chem. 275, 2269-2275 |
|
(3) |
Nakato, H., and Kimata K. (2002) Biochim. Biophys. Acta
1573, 312-318 |
|
(4) |
Yamada, S., Okada, Y., Ueno, M., Iwata, S., Deepa, S.S., Nishimura,
S., Fujita, M., Die, I.V., Hirabayashi, Y., and Sugahara, K. (2002)
J. Biol. Chem. 277, 31877-31886
|
|
(5) |
Toyoda, H., Kinoshita-Toyoda, A., Fox, B., and Selleck, S.B. (2000)
J. Biol. Chem. 275, 21856-21861
|
|
|