|
|
GAG惗崌惉僀僯僔僄僀僞堦 | |||||||||||||||||
 |
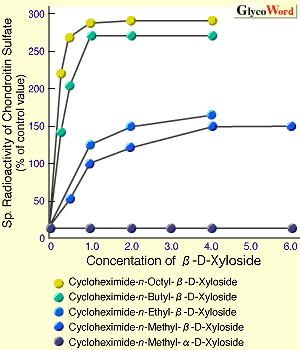
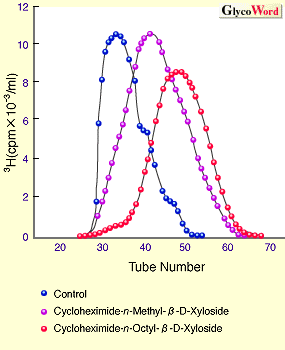
丂僾儘僥僆僌儕僇儞乮PG乯偼堦杮埲忋偺GAG偑僞儞僷僋恈偵嫟桳寢崌偟偨暋崌懱暘巕偱偁傞丅働儔僞儞棸巁埲奜偺棸巁壔GAG丄懄偪僐儞僪儘僀僠儞棸巁乮CS乯丄僨儖儅僞儞棸巁乮DS乯丄僿僷儔儞棸巁乮HS乯丄偍傛傃僿僷儕儞乮HP乯傪丄億儕儁僾僠僪乮PP乯偵寢崌偡傞嫶搉偟晹埵偼偡傋偰嫟捠偟偰偍傝丄GlcA兝1仺3Gal兝l仺3Gal兝l仺4Xyl兝1-0-PP峔憿傪桳偡傞丅偙傟傜偺GAG偺崌惉偼丄PP拞偺摿掕偺Ser偺悈巁婎偵UDP-Xyl偐傜Xyl偑揮堏偡傞偙偲偵傛傝奐巒偝傟傞丅GAG崌惉偵偍偄偰Xyl偑PP偵揮堏偡傞偙偲偲丄嫶搉偟晹埵偺旕娨尦枛抂偺GlcA偵GlcNAc偁傞偄偼GalNAc偑揮堏偡傞偙偲偲偼惗壔妛揑撪梕傪堎偵偡傞丅慜幰偼PP偵寢崌偡傞GAG嵔偺杮悢傪寛傔傞偨傔偺斀墳偱偁傝1 丄屻幰偼嫶搉偟晹埵偐傜怢挘偡傞GAG偺暘巕庬傪寛傔傞偨傔偺斀墳偱偁傞2 丅尵偄姺偊傟偽丄Xyl揮堏峺慺偑偳偺傛偆側忣曬傪幆暿偟偰Xyl傪揮堏偡傞偐偵傛傝寢崌偡傞GAG嵔偺杮悢偑寛傑傝丄偦偙偵宍惉偝傟偨嫶搉偟晹埵傪帩偮PP偑丄兛GlcNAcT1乮忋婰GlcA偵GlcNAc傪揮堏偡傞峺慺乯偁傞偄偼兝GalNAcT1(GlcA偵GalNAc傪揮堏偡傞峺慺)丄偄偢傟偺峺慺偺幆暿撪梕傪帩偮偐偵傛偭偰怢挘偡傞GAG偺暘巕庬偑寛傑傞丅峏偵,GAG怢挘奐巒場巕偺傾僌儕僐儞乮旕摐晹埵乯偑GAG偺嵔挿傪寛掕偡傞惈幙傪帩偮偙偲傪帵嵈偡傞寢壥傕摼傜傟偰偄傞3 丅埲壓丄忋偵徯夘偟偨撪梕偵廬偭偰丄侾乯Xyl揮堏峺慺偺帵偡幆暿撪梕丄俀乯兛GlcNAcT1偍傛傃兝GalNAcT1偺幆暿撪梕丄俁乯傾僌儕僐儞埶懚揑側GAG嵔挿偺寛掕偺條幃偵偮偄偰徯夘偡傞丅 侾乯Xyl偑揮堏偝傟傞摿掕偺Ser巆婎偼偳偺傛偆偵偟偰寛傔傜傟傞偺偐丠 丂cDNA偑僋儘乕僯儞僌偝傟偨PG僞儞僷僋恈偺傾儈僲巁攝楍傪斾妑専摙偡傞偙偲偵傛傝丄Xyl揮堏峺慺偺婎幙偲偟偰偺昁梫嵟彫儁僾僠僪扨埵偑悇掕偝傟偨丅偙偺儁僾僠僪撪偺偄偔偮偐偺傾儈僲巁傪懠偺傾儈僲巁偵姺偊偨曄堎懱偑崌惉偝傟丄Xyl揮堏斀墳偺庴梕懱偲偟偰丄奺乆偺儁僾僠僪偵偮偄偰丄Km抣丄Vmax抣偑媮傔傜傟偨丅Vmax/Km抣偐傜Xyl揮堏峺慺偵嵟傕揔偟偨傾儈僲巁攝楍偑寛掕偝傟丄偦偺峺慺偺婎幙擣幆晹埵偲悇掕偝傟偨丅偦偺寢壥偵婎偯偄偰僞儞僷僋恈偺cDNA偺曄堎懱偑嶌傜傟丄奺乆偺敪尰岠棪偑媮傔傜傟偨寢壥丄Xyl揮堏峺慺偑擣幆偡傞傾儈僲巁攝楍偲偟偰Glu/Asp-X-Ser-Gly乮倃偼擟堄偺傾儈僲巁乯偑採彞偝傟偰偄傞1 丅 俀乯GAG暘巕庬偼偳偺傛偆偵偟偰寛傑傞偐丠 丂偙偺栤戣偵摎偊傞幚尡偼丄摨堦僞儞僷僋恈忋偵CS媦傃HS嵔偺椉GAG嵔傪帩偮崿惉宆PG偱偁傞儀乕僞僌儕僇儞丄僔儞僨僇儞丄僌儕價僇儞傪儌僨儖暘巕偲偟偰師偺傛偆偵偟偰峴傢傟偨丅偙傟傜俁暘巕娫偵嫟捠偟偰尒弌偝傟傞GAG寢崌晹埵嬤朤偺嫟捠傾儈僲巁攝楍偵偄偔偮偐偺曄堎傪摫擖偟偨cDNA偑嶌惉偝傟丄CHO嵶朎偵摫擖屻丄敪尰偝傟偨GAG偺暘巕庬偲嶻惗岠棪偑夝愅偝傟偨2 丅偦偺寢壥丄HS嵔偺怢挘偵偼丄Ser-Gly偺C乗枛抂懁偵悢屄偺傾儈僲巁傪傊偩偰偰Glu傗Asp偐傜側傞巁惈傾儈僲巁偺僋儔僗僞乕偑懚嵼偡傞偙偲丄峏偵Ser-Gly偵椬愙偟偰Trp側偳偺慳悈惈傾儈僲巁偑懚嵼偡傞偙偲偑昁梫偱偁傞偙偲偑帵偝傟偨丅偙傟偵娭楢偟偰嫽枴偁傞偙偲偼丄GAG恖岺揑怢挘奐巒場巕偱偁傞兝-Xyloside偼HS嵔怢挘偺偨傔偵偼傾僌儕僐儞偑慳悈惈偱側偄偲HS嵔怢挘妶惈傪帵偝側偄偲偄偆偙偲偱偁傝4 丄慳悈惈傾儈僲巁偺梫媮惈偲椙偄堦抳傪帵偟偰偄傞丅埲忋偵摼傜傟偨寢壥傪傑偲傔傞偲丄HS嵔偺怢挘傪寛傔傞兛GlcNAcT1峺慺偺婎幙擣幆晹埵偼丄GlcA-Gal-Gal-Xyl偺嫶搉偟係摐晹埵丄Ser-Gly偵椬愙偟偨侾働偺慳悈惈傾儈僲巁丄偍傛傃Ser-Gly偺C乗枛抂懁偵懚嵼偡傞巁惈傾儈僲巁偐傜側傞僋儔僗僞乕偺俁晹埵偱偁傞偙偲偑帵嵈偝傟偰偄傞丅CS嵔怢挘傪寛掕偡傞兝GalNAcT1偺婎幙摿堎惈偵娭偟偰偼廩暘側忣曬偼摼傜傟偰偄側偄丅 俁乯GAG偺恖岺怢挘奐巒場巕兝-D-Xyloside偺傾僌儕僐儞埶懚揑側GAG偺嵔挿寛掕 丂GAG偺嵔挿寛掕偺婡峔偼枹偩柧傜偐偵偝傟偰偄側偄丅偟偐偟丄GAG偺嵔挿偑GAG怢挘懍搙偵傛偭偰寛掕偝傟偰偄傞偙偲傪帵嵈偡傞寢壥偑丄GAG嵔偺恖岺揑怢挘奐巒場巕偱偁傞兝-D-Xyloside傪梡偄傞偙偲偵傛傝摼傜傟偰偄傞3 丅恾侾偵兝-D-Xyloside偺傾僌儕僐儞晹埵傪悈梟惈壔崌暔偐傜慳悈惈壔崌暔偵曄壔偝偣丄擃崪慻怐偲曐壏偟丄兝-D-Xyloside偐傜CS嵔傪怢挘偝偣丄CS怢挘懍搙(恾1A)偲CS偺嵔挿乮恾1B乯傪夝愅偟偨寢壥傪帵偟偨丅偙偺寢壥偼丄傾僌儕僐儞偑慳悈惈偱偁傞傎偳CS嵔偺崌惉懍搙偑懍偔丄崌惉偝傟偨CS偺嵔挿偼抁偄偙偲傪帵偟偰偄傞丅偙偺偙偲偼丄GAG嵔怢挘偵偍偄偰傾僌儕僐儞晹暘偲丄GAG崌惉偺応偱偁傞僑儖僕懱枌偲偺憡屳嶌梡偼GAG嵔怢挘懍搙偺寛掕偵娭梌偟丄GAG偺嵔挿偼崌惉懍搙偵傛傝寛掕偝傟傞偙偲傪嫮偔帵嵈偟偰偄傞傕偺偲峫偊傜傟傞丅 | ||||||||||||||||
| |||||||||||||||||
| 壀嶳丂幚乮嫗搒嶻嬈戝妛丒岺妛晹乯 | |||||||||||||||||
| |||||||||||||||||
| 1998擭 6寧 15擔 | |||||||||||||||||
| |||||||||||||||||