|
|
海産無脊椎動物におけるレクチン |  | |||||||||||||
| (Update Issue: May.9, 2007) | ||||||||||||||
 | 海産無脊椎動物には海綿動物から原索動物まで多くの動物種にレクチンの存在が知られている。それらは分子量が1万程度のものから、数十万のものまで様々であるが、アミノ酸配列が明らかにされたものはそれほど多くはない。現在までにアミノ酸配列が明らかになっている海産無脊椎動物レクチンの多くはC型レクチンファミリーに(C型レクチンの多様性の項参照)属している。それらは高等脊椎動物のC型レクチンとは異なり、比較的分子量が小さく、一本のポリペプチド鎖がほぼ1個のC型糖認識ドメイン(carbohydrate recognition domain;CRD)から成っている(1,2)。しかしながら、糖結合活性については明らかではないが、カブトガニ(Tachypleus tridentatus)の体液凝固系C因子のように、C型CRDと、それ以外の複数個のドメインから形成されるタンパク質も見出されていることから(3)、同様に複数のドメインから成る海産無脊椎動物C型レクチンが存在することも考えられる。異なるレクチンのC型CRD間でのアミノ酸配列の相同性は比較的低く、近縁種間でも一般に40%以下である。CRDには1カ所の糖結合部位が存在するが、海産無脊椎動物にはガラクトースやN-アセチルガラクトサミンに対する結合特異性を示すものが多い。一方、C型レクチン以外にも、海産無脊椎動物中にはこれまで分類された動物レクチンファミリーのいずれにも含まれないものもいくつか見出されている。例えば、カブトガニのリポ多糖結合性のレクチンは約33-38残基のアミノ酸配列の6回繰り返し構造を有している。また、ムラサキウニ(Anthocidaris crassispina)の卵から得られたガラクトース特異的レクチンについても、これまでに知られているどのタンパク質のアミノ酸配列とも相同性が見られないことが明らかになっている。 | |||||||||||||
| ||||||||||||||
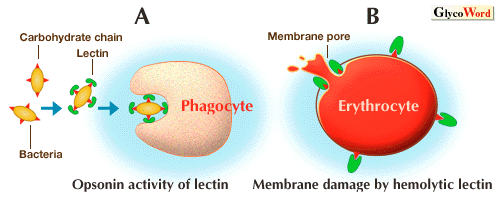
| 海産無脊椎動物レクチンの主な機能としては高等脊椎動物における免疫グロブリンに相当する生体防御因子としての役割が挙げられる。このことは、外来異物表面糖鎖に結合することによって食細胞を活性化(オプソニン作用)するレクチンが存在すること、また動物体内に異物を接種した際に、その体液中のレクチン活性が上昇することなどから示唆されている。一方、レクチン単独で細胞傷害性を示す例も最近明らかになった。カブトガニやナマコの一種であるグミ(Cucumaria echinata)からは、それぞれシアル酸やガラクトース特異的な溶血活性をもつレクチンが精製されているが、これらは赤血球表面へ糖鎖を介して結合した後に、細胞膜に直接傷害を与えることによって溶血作用を現すものと思われる。これらについては外部から侵入した微生物 に対する防御や、捕食者から身を守るための毒素としての役割が考えられる。一方、ラッパウニ(Toxopneustes pileolus)の叉棘(さきょく;とげの一種)毒中にはマイトジェン活性や遊走活性のような生理活性を示すレクチンの存在が認められており、これも毒作用との関連が示唆されている。その他にもいくつかのC型レクチンにおいて異なった機能が示されている。例えば、アカフジツボ(Megabalanus rosa)レクチンとミサキマメイタボヤ(Polyandrocarpa misakiensis)レクチンは、それぞれ炭酸カルシウム結晶化調節や形態形成へ関与していることが示唆されている。また、ムラサキイガイ(Mytilus edulis)精子レクチンは卵膜溶解因子として機能することが明らかにされている。このように、海産無脊椎動物レクチンは生体防御因子としての働き以外にも、様々な組織や体液中において、脊椎動物と同様、多くの異なった機能を担っていることが予想される。 | ||||||||||||||
| 畠山 智充(長崎大学・工学部) | ||||||||||||||
| ||||||||||||||
| 1998年 3月 15日 | ||||||||||||||
| ||||||||||||||