

|
|
リピド中間体生合成段階に変異を持つヒト遺伝病 |
|||||||||||||||||||||||||||||
 |
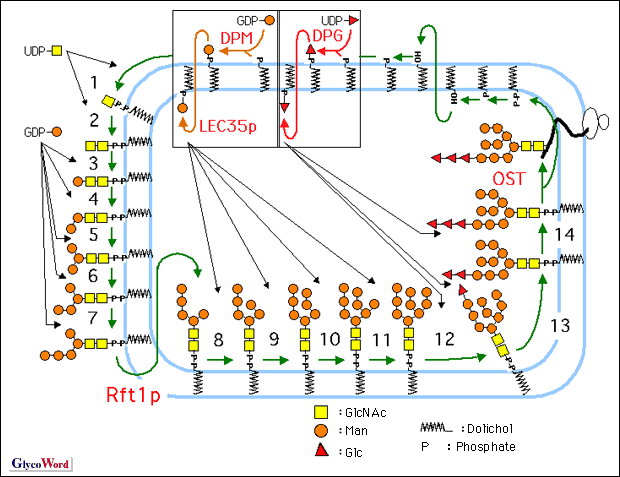
リピド中間体[Glc3Man9GlcNAc2-P-P-ドリコール(Dol)、下図]は糖蛋白のアスパラギン結合型糖鎖の前駆体である。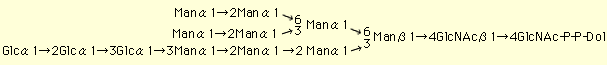 リピド中間体の生合成は粗面小胞体膜に存在する14種類の糖転移酵素によって行われる(Fig. 1: ステップ1から14)。まず、前半は、粗面小胞体膜の細胞質側で、糖ヌクレオチド(UDP-GlcNAcおよびGDP-Man)を糖供与体として、Man5GlcNAc2-P-P-Dolまで生合成された(Fig. 1: ステップ1から7)のち、内腔側に転送され、続く後半は、ドリコールリン酸糖(Dol-P-ManおよびDol-P-Glc)を糖供与体としてGlc3Man9GlcNAc2-P-P-Dolまで生合成されて完成する(Fig. 1: ステップ8から14)。この過程は真核細胞において高度に保存されている(1)。 この生合成には、キャリヤー・リピドであるドリコールリン酸(Dol-P)のde novo合成およびリサイクリングによる供給(2)、糖供与体である糖ヌクレオチドやドリコールリン酸糖の供給、14種類の糖転移酵素、糖受容体(未完成のリピド中間体)が関わるほか、永年、仮想されてきて、ごく最近、その手がかりが報告された、Man5GlcNAc2-P-P-Dolおよびドリコールリン酸糖の転送蛋白(flippase)[Man5GlcNAc2-P-P-Dol転送蛋白(Rft1蛋白(Fig. 1、3))、Dol-P-Man転送蛋白(LEC35蛋白)( Fig.1、4)およびDol-P-Glc転送蛋白(LEC35様蛋白)]などが関わっている。そして、それらは更に転写の段階や酵素反応の段階などで、様々な調節制御を受けている。 複合糖質の糖鎖生合成に関する遺伝病に先天性グリコシル化異常症[Congenital Disorders of Glycosylation: CDG]がある。CDGは1980年にJaekenらによって、重篤な脳神経症状をもたらし、かつ、生化学的解析による血清糖蛋白トランスフェリンの等電点電気泳動における易動度の異常(シアル酸量の減少に基づく)を示す遺伝病としてはじめて報告されたものである。現在では、CDGはtype I (CDG-I)とtype II (CDG-II)に大別されている。生化学的には、CDG-Iはタンパクあたりのアスパラギン結合型糖鎖の数の減少として検出されるものであり、CDG-IIはアスパラギン結合型糖鎖のプロセシングの異常として検出されるものである(5)。 このうち、CDG-Iの病因としては、リピド中間体の供給減少とオリゴ糖転移酵素(Fig.1: OST)の活性低下をもたらす様々な段階の変異が考えられよう。そして、現在までに、以下に示すような7種類のCDG-Iのsubtypeが見つかっている。すなわち、糖リン酸生合成段階の変異したものとしてホスホマンノムターゼ(Man-6-PからMan-1-Pを生成)遺伝子(PMM2)の変異(CDG-Ia、現在までに、約350症例)やホスホマンノースイソメラーゼ(Fru-6-PからMan-6-Pを生成)遺伝子(PMI)の変異(CDG-Ib、現在までに約30症例)が知られている。これらの患者では、結果としてGDP-ManおよびDol-P-Manが欠乏し、リピド中間体の供給が減少することがunderglycosylationをもたらす。このうち、CDG-Ibの場合はマンノースの経口投与により治療可能である。更に、近年、リピド中間体生合成段階に変異があるためにunderglycosylationにいたる患者も、それぞれ、1~12症例見つかっている。すなわち、グルコース転移酵素I(Fig. 1: ステップ12)の遺伝子(hALG6)の変異の結果、Man9GlcNAc2-P-P-Dolを蓄積するCDG-Ic、マンノース転移酵素VI(Fig. 1:ステップ8)の遺伝子(hALG3)の変異の結果、Man5GlcNAc2-P-P-Dolを蓄積するCDG-Id、Dol-P-Man合成酵素(Fig. 1:DPM)のDpm-1サブユニットの遺伝子(hDPM1)の変異の結果、Man5GlcNAc2-P-P-Dolを蓄積するCDG-Ie、Dol-P-Manの粗面小胞体膜における転送に働くと考えられているLEC35蛋白(Fig. 1: LEC35p)の遺伝子(MPDU-1/LEC35)の変異の結果、Man5GlcNAc2-P-P-Dol、Man9GlcNAc2-P-P-Dolなどを蓄積するCDG-If、マンノース転移酵素VIII(Fig. 1:ステップ10)の遺伝子(hALG12)の変異の結果、Man7GlcNAc2-P-P-Dolを蓄積するCDG-Igである。現在までの所、リピド中間体生合成過程の前半に欠陥を持つCDG-Iは見つかっていない。 リピド中間体生合成に関わる遺伝子は出芽酵母を中心として分離されてきた(1)が、まだ、2、3の遺伝子はクローニングされていない。また、出芽酵母のリピド中間体生合成関連糖転移酵素遺伝子をもとにヒト相同遺伝子がクローニングされている。しかし、ヒトには、出芽酵母とは異なる構成をもつ酵素・遺伝子や出芽酵母に見いだされていない遺伝子も存在する。筆者らはGDP-Manをマンノース供与体とするマンノース転移酵素群に焦点をあて、マンノース転移酵素Iのヒト遺伝子(Hmat-1)などのホモロジー・クローニングと新規マンノース転移酵素遺伝子の発現クローニングを行っている(6-8)。 さきに述べたように、リピド中間体生合成は複雑な過程であり、その解明と平行して、これからも、直接そして間接に、リピド中間体生合成に関わるCDG-Iの新しいsubtypeが見つかるものと思われる。 |
||||||||||||||||||||||||||||
 |
|||||||||||||||||||||||||||||
| Fig.1 リピド中間体生合成サイクルとそのトポロジー | |||||||||||||||||||||||||||||
| 高橋哲夫・西川義尚 (東海大学工学部・生命化学) | |||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||
| 2002年 6月 15日 | |||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||

