

|
|
O-GlcNAcと細胞機能 |
||||||||||||||||||||||||||||||||
 |
核および細胞質内に存在する種々のタンパク質には,セリンあるいはスレオニン残基の水酸基の酸素に, |
|||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
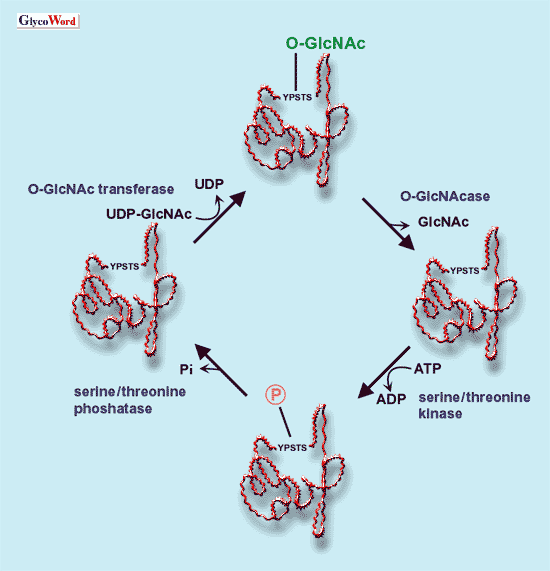
| 成熟ラット膵臓でO-GlcNAcの発現を免疫組織化学的に解析すると、O-GlcNAcはランゲルハンス島の内分泌細胞の核と細胞質に見い出され、とりわけ核に多量に存在した(2)。現在まで、核および細胞質に存在するO-GlcNAcをもつタンパク質が多数同定されている。主なものとしては、転写因子、核膜孔タンパク質、ガン遺伝子産物、ガン抑制因子、細胞骨格タンパク質である。これらのタンパク質に共通していることは、?リン酸化タンパク質であること、?他のタンパク質と可逆的に複合体を形成するという、2点である。これらのことから、O-GlcNAc
修飾はタンパク質のリン酸化を調節することによってシグナル伝達に関与し,タンパク質の会合やその安定化に寄与すると考えられている。 O-GlcNAcの修飾(付加と除去)に関与する酵素が、精製されクローニングされている(3, 4)。O-GlcNAc転移酵素(UDP-GlcNAc:polypeptide O- 今日では、O-GlcNAcの修飾の異常が、糖尿病,癌、アルツハイマー病などの疾患と密接に関連していることが明らかにされつつある(1, 6-9)。種々の組織で、O-GlcNAcが担う細胞機能の詳細な解明が待たれる。 |
||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
| 秋元 義弘(杏林大学医学部 解剖学第2講座) | ||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
| 2003年 1月22日 | ||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||

