

|
|
ガングリオシドの糖鎖構造に依存したエクト型NAD分解酵素CD38の阻害 |
 |
|||||||||||||
 |
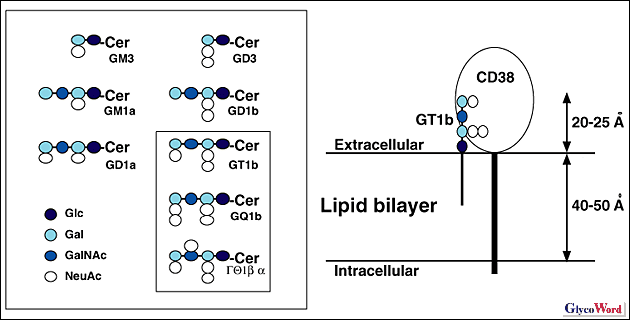
ガングリオシドはシアル酸を含む糖脂質であり、細胞膜の表層側に多く存在する。細胞上のガングリオシド糖鎖は別の細胞の表面のタンパク質により認識されることが可能であるが、同一細胞の表面のタンパク質により認識されることもまた可能である。細胞が糖鎖を脂質に結合して表面に提示する生理的な意味として、どちらの種類の認識がより重要であるのかは現在未解決の問題である。ガングリオシド糖鎖と同一細胞上のタンパク質の相互作用の例として本稿ではガングリオシドの糖鎖構造に依存した細胞表面上酵素の阻害の現象を紹介したい。 細胞外ドメインに触媒活性をもつ酵素をエクト型酵素とよぶ。リンパ球表面抗原CD38は分子量46 kDaの1回膜貫通型タンパク質であり、細胞外ドメインにNAD分解活性をもつエクト型酵素である。CD38の遺伝子を欠損したマウスでは全身の組織のNAD分解活性がほぼ完全に消失するのでCD38は動物細胞における主要なNAD分解酵素である。CD38の細胞外ドメインのNAD分解活性は種々のガングリオシドにより阻害される。阻害の程度は糖鎖構造に依存し、特に内側のグルコースにタンデムシアル酸が結合したb系列ガングリオシド(GT1b, GQ1b, GQ1b CD38を発現している細胞にGT1bを取り込ませても阻害が起こる。阻害のタイムコースはGT1bの取り込みのタイムコースよりも遅いので、GT1bが細胞表面にとりこまれてから阻害が起こる。阻害はGT1bとCD38が同一細胞表面にあるときに効率良く起こる。またb系列ガングリオシドGT1bとこれよりひとつだけシアル酸が少ないa系列ガングリオシドGD1aを比較すると明らかにGT1bの阻害効果の方が大きい。これらの結果からGT1bの糖鎖が同じ細胞表面のCD38と相互作用することが考えられる (3)。 ガングリオシドの糖鎖は目一杯伸びた構造をとっても細胞膜表面(脂質の非極性部分と極性部分とのつなぎ目)から20-25オングストローム以内の範囲に存在する。タンパク質に結合した糖鎖に比較してはるかに膜からの距離が短い。ガングリオシド糖鎖を認識するタンパク質の道具立てを明らかにすることは、細胞が膜表面のごく近くに糖鎖を提示する生理的意義を解明するために重要である。 |
|||||||||||||
 |
||||||||||||||
| Fig. 1 | ||||||||||||||
| 横山三紀 (東京医科歯科大学医歯学総合研究科・硬組織病態生化学) |
||||||||||||||
|
||||||||||||||
| 2003年 1月22日 | ||||||||||||||
|
|
||||||||||||||
|
||||||||||||||

