

|
|
GM3合成酵素 | ||||||||||||||||||||
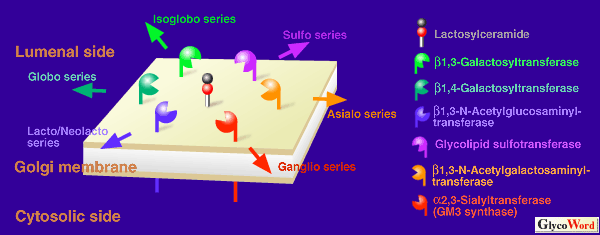
 | ガングリオシドGM3は生体内主要ガングリオ系列ガングリオシドの共通の前駆体で、ほぼ全ての哺乳動物組織中に存在する。GM3は、CMP-シアル酸由来のシアル酸をラクトシルセラミドの非還元末端ガラクトースにα-2,3結合で転移することで生合成されるが、この反応を触媒するのがGM3合成酵素(CMP-シアル酸:ラクトシルセラミドα-2,3シアル酸転移酵素;SAT-I:EC. 2.4.99.9)である。本酵素は生体内糖脂質糖鎖生合成の分岐点で作用するので、その活性発現調節はガングリオ系のみならず、ラクト・ネオラクト系、グロボ系、イソグロボ系などの糖脂質生合成にも影響を与えるものと思われる。 | |||||||||||||||||||
| ||||||||||||||||||||
|
1991年、Melkerson-Watsonらは、ラット肝より4万3千倍まで精製し(1)、また1993年にはPreussらが、ラット脳より7千2百倍に精製した(2)。両者には分子量や基質特異性などの点で相違があり、同一のタンパク質かどうか定かでない。後者に関しては、酵素活性調節にリン酸化/脱リン酸化が関与しているとの報告がある(3)。 GM3がもつ生体機能に分化誘導作用がある。HL-60細胞をホルボールエステルで処理すると単球様細胞へ分化する。この際SAT-I活性の上昇に伴い、細胞内GM3量が著増する。更に、GM3をHL-60細胞に添加すると、ホルボールエステル処理時と同様に単球様細胞へ分化した(4)。1997年、この系を利用し、発現クローニング法によりSAT-I遺伝子がクローニングされた(5)。SAT-Iは、シアル酸転移酵素ファミリー(STs)に属する新規のタンパク質であったが、シアリルモチーフL(シアリルモチーフの項参照)中で、他のSTsでは保存されているアスパラギン酸がヒスチジンに置換しているという構造的特徴を示した。このアミノ酸置換は、マウス・ラット由来のSAT-Iにおいて共通に見られた。 糖脂質をシアル酸受容体とする他のSTsと異なり、SAT-I遺伝子産物の基質特異性はラクトシルセラミドに限局されていた。この点は、上記精製SAT-Iの結果とも相違する。また、GM3の汎在性にもかかわらず、SAT-ImRNAの発現には、組織特異性並びに動物種特異性が認められた。 遺伝子クローニングにより、GM3の生体機能解明に拍車がかかることが期待される。また、SAT-I遺伝子を手がかりに、動物界におけるGM3の起源をたどる試みは、GM3の生物学的重要性を示す一つの試みとして興味がもたれる。 | ||||||||||||||||||||

| ||||||||||||||||||||
| 石井 睦 斎藤 政樹(国立がんセンター研究所・ウイルス部) | ||||||||||||||||||||
| ||||||||||||||||||||
| 1998年 12月 15日 | ||||||||||||||||||||
| ||||||||||||||||||||

