|
|
糖脂質糖鎖―糖鎖相互作用 | |||||||||||||||||
 |
ガン化や発生に伴う劇的な細胞表層糖鎖の変化が示されて以来、細胞間や細胞と基質間の認識における糖鎖の役割について議論され、糖鎖に基づく細胞間相互作用に関する多くのモデルが提唱されている。その多くにおいて糖鎖はセレクチンやガレクチンあるいはその他の糖鎖結合性たんぱく質と糖鎖ーたんぱく質間相互作用によって結合するとされている。一方、Hakomoriらは糖鎖に依存する細胞接着の新しい機構を提出している。すなわち、糖鎖ー糖鎖間の相互作用によって細胞間相互作用の始まるというモデルである。 糖鎖―糖鎖間相互作用の最初の証拠は高度に精製した糖脂質を含むリポソームを用いた研究によってもたらされた。以下に述べるような実験事実は特異的な糖脂質がそれと相補的な糖脂質に糖脂質の糖鎖部分による相互作用を介して結合することを示唆している。
図1にこれまでに観察された可能な糖脂質―糖脂質相互作用をまとめた。例えばマウス初期胚のコンパクションのモデルとしてLex-Lex 間の相互作用がHakomori らにより見い出された。Lexを発現している細胞、あるいは外部からLex-糖脂質を取り込ませた細胞では2価イオン存在下で凝集する。GM3という糖脂質を非常に強く発現しているマウスメラノーマB16細胞はGg3Cerと非常に強くLacCerと強くまたGb4Cerと弱く結合するが、この結合様式はGM3を含むリポソームを用いた場合と全く同じである。またB16細胞はGg3Cerを発現している L5178 AA12細胞やLacCerを発現している血管内皮細胞に接着する。このこれらの接着や凝集は相当する糖脂質を含むリポソーム、あるいは糖脂質糖鎖と相同なオリゴ糖、相当する糖脂質を認識する抗体によって阻害することができるが、糖脂質を含まないリポソームや他の抗体、オリゴ糖などでは阻害されない。 | ||||||||||||||||
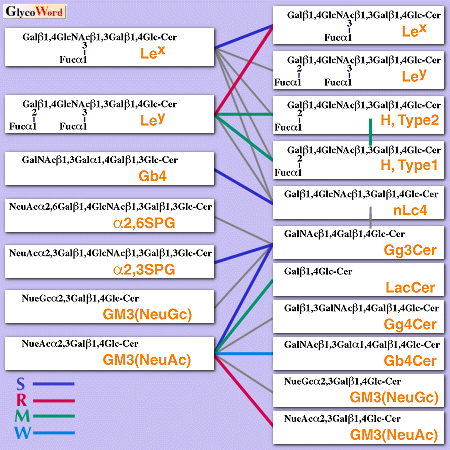 | |||||||||||||||||
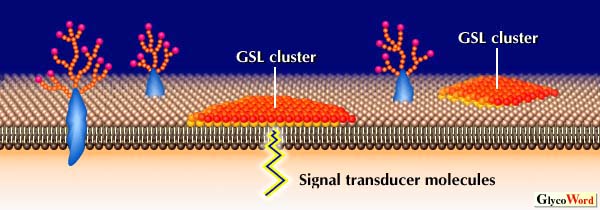
| 図 1 Possible GSL-GSL interaction observed using GSL liposomes. S, M, W,and R indicate strong, moderate, weak and repulsive interaction, respectively. Gray line indicates no interaction. | |||||||||||||||||
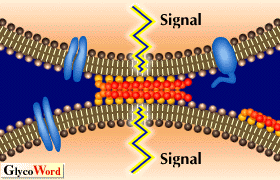
| 図1に示したいくつかの糖脂質―糖脂質相互作用については分子間力を測定すること、あるいはイオンスプレーマススペクトルによっても確かめられている。しかしNMRを用いた糖脂質の糖鎖部分の直接的な相互作用についてはまだ示されていない。糖脂質を含むリポソームの糖脂質を固相化したプレートへの結合が糖脂質の密度に依存することから、糖脂質ー糖脂質間の相互作用には糖脂質の多価化あるいはクラスター化が必要であると考えられる。Tillackらは電子顕微鏡を用いた研究から糖脂質が細胞表面やリポソーム上でクラスターとして存在していることを示している。また糖脂質とリン脂質の単相膜上での相分離についてもsurface pressure-area isothermsを測定することで確認されている。しいたがって細胞表面の糖脂質はおそらく大きなクラスターとして存在し細胞表面上でミクロドメインを形成していると考えられる(図2)。このような糖脂質クラスターは抗体やレクチンだけでなく、相補的な糖脂質のクラスターとも強く結合できるかも知れない。細胞や糖脂質を含むリポソームの糖脂質を固相化したプレートへの特異的接着はこのような糖脂質クラスター間の相互作用によるものと考えられる。また糖脂質―糖脂質相互作用はたんぱく間の相互作用より弱いが素早くおこる。さらにインテグリンなどの接着分子による細胞接着と糖脂質ー糖脂質相互作用による細胞接着の間に共同関係があることも見出されている。従って、糖脂質―糖脂質間の比較的弱い相互作用は細胞認識の初期の特異性や方向性を決定し、それに続く様々な接着過程を調節しているのかも知れない。 | |||||||||||||||||
| |||||||||||||||||
| ある糖脂質を強く発現している細胞はそれに相補的な糖脂質を固相化したプレート上に強く結合するだけではなく、そのうえで細胞の伸長や運動が促進されることが見い出されている。最近糖脂質からなるミクロドメインにはC-Src, Ras, Lyn, FAKといったシグナル伝達分子が高度に含まれていることが示された。また、細胞中でのFAKのチロシンリン酸化等が相補的な糖脂質を固相化したプレートに細胞が接着した場合に促進されることも報告されている。従って糖脂質のクラスターは細胞接着の場として働くだけでなく、情報伝達の場としても機能しているのかも知れない。未だに糖脂質ー糖脂質相互作用が生体内でおこっているという証拠はないが、糖脂質ー糖脂質相互作用が細胞間の認識や情報伝達においてなんらかの役割をになっている可能性は否定できない。 | |||||||||||||||||
| 小島 直也 (東海大学・工学部) | |||||||||||||||||
| |||||||||||||||||
| 1998年 9月 15日 | |||||||||||||||||
| |||||||||||||||||