

|
|
糖脂質マイクロドメイン |  | |||||||||||||
| (First version published: 1998年6月15日) | ||||||||||||||
 | スフィンゴ糖脂質は飽和アシル鎖を持つものが多く、細胞膜の主要成分である不飽和アシル鎖に富むリン脂質から相分離することにより、ドメイン構造を取ると考えられている。グリセロ糖脂質においても、ステアリン酸(C18:0)とアラキジン酸(C20:0)の飽和アシル鎖をそれぞれ、C1位とC2位に有するホスファチジルグルコースは、HL60細胞でドメインを形成する。このようなマイクロドメインに情報分子が集合することで、膜分子同士の会合の確率が上がり、膜を介した情報伝達の効率を上げている。 | |||||||||||||
 |
||||||||||||||
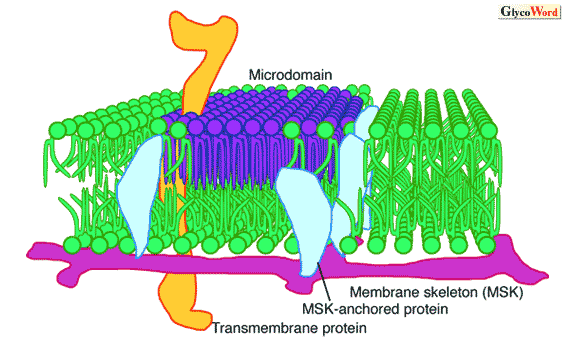
| 図 膜骨格アクチンとそれに繋がれたタンパク質の”ピケット”により動きが制限された膜脂質とマイクロドメイン | ||||||||||||||
|
当初、raftやDIGと称された糖脂質に富むマイクロドメインはコレステロールに富むとされていたが、そうではないドメインも存在する。箱守らはガングリオシドに富むマイクロドメインで、コレステロールをほとんど含まず、細胞接着や細胞間認識に関わるものを「グリコシナプス」と称して他と区別している。ここでは、CD9, CD81等の4回膜貫通タンパク質であるテトラスパニン、インテグリン受容体、細胞増殖因子受容体等がガングリオシドと会合し、それらの相互作用がインテグリンや細胞増殖因子のシグナルを調節することで、細胞の形質の転換を調節する。raft等と同様、この膜ドメインの細胞質側にはSrcファミリーキナーゼや小型Gタンパク質が集まる。細胞内外の二重膜にまたがって存在する受容体タンパク質やテトラスパニン等が、これらシグナル分子の膜の両側への集合を橋渡ししているのであろう。 生理的な温度では細胞膜を形成する脂質は液体として常に運動しているので、そこに浮かぶ膜分子も常に動き廻っている。したがって、raftやグリコシナプス等のマイクロドメインは、常に変動していると考えられる。その動きは、アクチンからなる細胞膜の内張タンパク質によって形成される膜骨格の編み目によって制限される。その大きさは細胞によって異なるが、30-230 nm程度である。この区画内においては、膜タンパク質や脂質は熱運動による自由な動きが可能であるが、膜貫通型タンパク質は、この膜骨格に引っかかることでその動きが制限される(フェンスモデル)。また、膜骨格に繋がっている膜タンパク質は、膜脂質の区画外への自由な移動を妨げる(ピケットモデル)。その結果、膜分子は、一つの区画内にしばらく留まった後、隣の区画に移動する。マイクロドメインは、この膜区画内に形成される。マイクロドメインにおける情報伝達には、膜区画の存在が影響を及ぼしている。リガンドが結合した受容体などが膜骨格に一時的に結合することによって、細胞への入力の位置情報を与えていると考えられている。 |
||||||||||||||
| 東 秀好(東北薬科大学分子生体膜研究所 生体膜情報学研究室) | ||||||||||||||
| ||||||||||||||
| 2007年 2月 1日 | ||||||||||||||
| ||||||||||||||

