

|
|
β-1,4-ガラクトース転移酵素(β-1,4-GalT)遺伝子KOマウス |
 |
|||||||||||||||||||||||||
 |
β-1,4-ガラクトース転移酵素(β-1,4-GalT)(EC 2.4.1.38)は,2型基幹構造(Galβ1→4GlcNAc)の生合成に必須な糖転移酵素であり,この構造は糖タンパク質のN型糖鎖やO型糖鎖並びに糖脂質に広く見られる。特に,2型基幹構造は免疫系で重要な役割を担っているシアリルルイスxや初期発生に重要なSSEA-1に必須である。β-1,4-GalT遺伝子は最近までひとつと思われてきたが,1997から1998年に次々と新しい遺伝子が単離され,現在7つからなる遺伝子ファミリーを形成していることが明かとなった。最初に単離されたβ-1,4-GalT-IのKOマウスが,我々とShurらにより作製された(1, 2)。β-1,4-GalT-IのKOマウスにおいて,若干のβ-1,4-GalT残存活性や2型基幹構造に対応する薄いバンドが検出されたが,これらは他のβ-1,4-GalT遺伝子によるものだと考えられる。 | |||||||||||||||||||||||||
 |
||||||||||||||||||||||||||
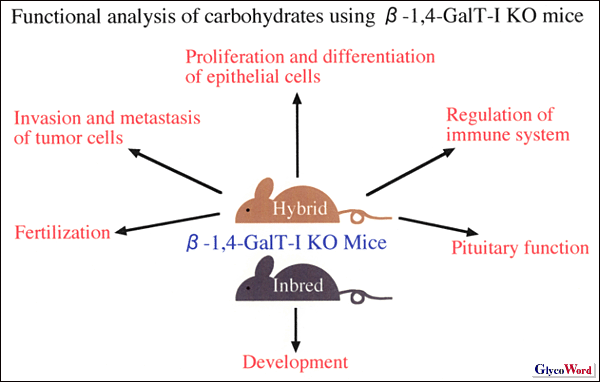
| Galβ1→4GlcNAc構造は初期発生での重要性が示唆されていたが,β-1,4-GalT-I KOマウスは,予想に反して正常に生まれてきた。しかし,出生後に成長遅延を示し,離乳までに約半数が死亡した。Shurらは脳下垂体ホルモンにガラクトースが付加されないことによる脳下垂体機能不全が原因であるとしている(2)。一方,我々は小腸絨毛細胞の分化異常,特にラクターゼの発現低下が成長遅延と致死の原因と考えている(1)。β-1,4-GalT-I KOマウスにおいて皮膚や小腸の上皮細胞の増殖亢進と分化異常が見られ,β-1,4-GalT-Iの合成する糖鎖は上皮細胞の増殖と分化を制御していることが示唆された(1)。 Shurらのこれまでの研究では,精子の細胞表面に存在する長いタイプのβ-1,4-GalT-Iが卵子のZP3と結合して,受精に関与することが示唆されていたが,β-1,4-GalT-I KOマウスは自然交配では雌雄とも妊孕性に問題がなかった。そこでShurらは体外受精の系で解析したところ,β-1,4-GalT-I KO精子は,ZP3に対する結合が低下しており,卵への侵入効率が悪いことを報告している(3)。このことは,自然交配では精子と卵子の結合に関与する他の分子が,β-1,4-GalT-I欠損を相補している可能性を示唆している。 β-1,4-GalT-I KOマウスを用いて,β-1,4-GalT-I欠損が様々な組織におけるN型やO型糖鎖に与える影響を解析した。Galβ1→4GlcNAcの外側に形成されるポリシアル酸やHNK-1などの糖鎖は,神経系の構造と機能に重要であるが,β-1,4-GalT-I KOマウスの行動は正常であり,脳でのこれらの糖鎖構造の発現も正常であった(4)。脳においてはβ-1,4-GalT残存活性が野生型の65%もあり,他のβ-1,4-GalT遺伝子が働いていることが示された。赤血球膜上の糖タンパク質糖鎖の構造を解析したところ,β-1,4-GalT-Iは,赤血球においてO型糖鎖のcore 2の生合成を担っていることがわかった(5)。更に,肝臓細胞膜や血漿糖タンパク質について,N型糖鎖構造を詳細に解析したところ,β-1,4-GalT-I KOマウスは,2型基幹構造(Galβ1→4GlcNAc)が合成されないだけでなく,1型基幹構造(Galβ1→3GlcNAc)の合成が促進されていることがわかった。また,N型糖鎖の基幹構造の変化に伴い,末端のシアル酸もSiaα2→6GalからSiaα2→3Galへの変換が認められた。以上のことからβ-1,4-GalT-Iの欠損は,単純に2型基幹構造が欠損するのではなく,基幹糖鎖構造の大きな変化をもたらしていることが明かとなった (6)。このことはこのマウスの表現型の解釈に重要である。 β-1,4-GalT-I KOマウスのその後の解析から,白血球でもO型糖鎖のcore 2の合成が低下しており,白血球へのセレクチンの結合が低下して,その結果,炎症反応が低下していることがわかってきた。また,近交系化するとβ-1,4-GalT-I KOマウスは胎生致死を示すようになり,発生過程にも重要な役割を果たしていることが明かとなりつつある。 ごく最近,β-1,4-GalT-I 遺伝子に突然変異の入ったヒトの患者が見つかり,CDG-IIdと命名された(7)。C末端側が50アミノ酸欠損したβ-1,4-GalT-Iタンパク質を発現しているが,ゴルジ体には局在しない。β-1,4-GalT-I残存活性がβ-1,4-GalT-I KOマウスと同じ程度(5%)であるので,完全な欠損型と考えられる。しかし,患者は,水頭症や筋障害,血液凝固異常を示し,β-1,4-GalT-I KOマウスの症状とは一致しない。ヒトの患者はたった一例なので,今後マウスとヒトの比較をはじめとした詳細な解析が必要である。 |
||||||||||||||||||||||||||
| 浅野雅秀(金沢大学大学院医学系研究科・附属動物実験施設) | ||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
| 2002年 6月 15日 | ||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||
|
||||||||||||||||||||||||||

