

|
|
Nodファクターの認識・伝達機構 |
||||||||||||||||||||
 |
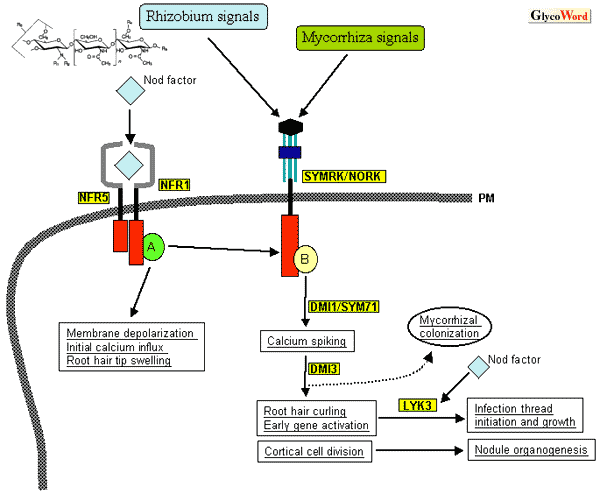
窒素固定共生をもたらすマメ科植物と根粒菌の相互作用は、根粒菌の生産するアシル化キチンオリゴマー(Nodファクター)によって開始される。Nodファクターの基本的な構造やその作用については先に紹介した(Nod-factor を介したマメ科植物・根粒菌の相互作用:糖質科学のことば、1999)ので、ここではその後の研究の重要な進歩に焦点を合わせて解説したい。 Nodファクターを宿主マメ科植物に投与すると、まず根毛細胞の脱分極、細胞外pHの上昇、Caイオンの一過的な流入とそれに引き続く流出入(Caスパイキング)が引き起こされる。これらと前後して根毛の先端肥大、変形、枝分かれなど根粒菌感染に特徴的な現象が観察される。Nodファクターはまた、皮層細胞の分裂を誘導し根粒原基を形成させる。こうした一連の反応は、根の表層できわめて局所的に起こるため、その生化学的な解析は容易でなく、Nodファクターの構造が1990年に初めて決定されて以来、マメ科植物によるNodファクター受容系は長い間謎に包まれていた。 しかし、ミヤコグサ(Lotus japonicus)、タルウマゴヤシ(Medicago truncatula)などのモデルマメ科植物を用いた最近の分子遺伝学的研究によって、Nodファクター受容体を含め、初期のシグナル応答・伝達系に関与する多くの遺伝子がクローニングされるようになった。受容体候補遺伝子は、ミヤコグサの根粒形成欠損(nod-)変異体Ljsym1、Ljsym5からポジショナルクローニングにより単離された。これらの変異体は、Nodファクターに対して上述のような初期応答のすべてを欠損していることから、その原因遺伝子はNodファクター受容体そのものあるいはその直下のシグナル伝達系に位置づけられるものと推定されていた。変異の原因遺伝子はそれぞれNFR1、NFR5と名付けられ、ともにLysMドメイン-膜貫通領域-キナーゼドメインよりなるレセプターキナーゼをコードしていた。LysMは細菌のペプチドグリカン分解酵素の基質認識に関与するドメインで、ペプチドグリカンの基本骨格がN-アセチルグルコサミンとN-アセチルムラミン酸がβ-1,4結合した構造を持つことから、単離された遺伝子の産物は細胞膜外に存在するLysMドメインによってNodファクターを認識し、細胞内のキナーゼドメインによってそのシグナルを次のステップに伝達していると予想される。 一方、NFR1/5の発見にわずかに先立って報告されたタルウマゴヤシのLYK3も、LysMファミリーに属するレセプターキナーゼであり、根粒菌のstrain-specificな認識に関与するlocusとして古くから知られていたエンドウSYM2のオルソログとして単離された。LYK3/SYM2は、ある特定の化学修飾をもつNodファクターを認識して感染プロセスを進行させるのに必須の役割を果たしている。実際、RNAiによるLYK3のノックダウンによって、感染糸形成はおこるもののその伸長が初期に停止してnod-となることが示された。さらに、この感染糸の生長停止は、Nodファクターの化学構造に依存していた。根粒菌は感染糸を形成しつつ宿主植物細胞中に侵入する過程でもNodファクターを生産し続けており、感染プロセスの進行においてもLysM型レセプターキナーゼによる特異的なNodファクター認識が必須であると考えられる。 ところで、植物と微生物とのより普遍的な共生系であり、植物へのリン酸供給に重要な役割を果たしている菌根菌(mycorrhiza)共生は、マメ科のみならずほとんどの植物に広く分布し、進化的には根粒菌共生よりも遙か以前に成立していたと推定される。nod-変異系統の半分以上は同時に菌根菌共生能も欠損している(myc-)ことから、これら2つの共生系は一部共通する遺伝的基盤の上に成り立っていると考えられる。NFR1/NFR5変異体は菌根菌共生は正常(myc+)であり、そのこと自体、これらの変異がNodファクター受容系そのものの破綻であることを強く示唆している。しかし、nod-myc-変異体の多くは、Nodファクターに対して全く無応答というわけではない。nod-myc-ミヤコグサ変異体からクローニングされたSYMRK(タルウマゴヤシではNORK)は、ロイシンリッチリピート(LRR)を膜外ドメインにもつ、やはり膜貫通型レセプターキナーゼである。SYMRK変異体は、Nodファクターに対して野生型と同様な根毛の変形を示すが、Caスパイキングは見られず、またレグヘモグロビンなど共生に特異的な遺伝子発現を誘導することもない。そのため、SYMRKはNodファクターの受容からCaスパイクの誘導に至る相互作用初期のシグナル伝達経路に含まれると推定されている。さらにDMI1(タルウマゴヤシ)/SYM71(ミヤコグサ)、DMI3(タルウマゴヤシ)など、Nodファクター初期シグナル伝達系に位置づけられる遺伝子がごく最近相次いでクローニングされた。 興味深いことに、タルウマゴヤシのLYK3遺伝子の近傍ゲノム領域には非常に多くの類似のレセプターキナーゼが存在していた。ミヤコグサのNFR1についても同様で、Nodファクター受容体(候補)遺伝子群は、ゲノム上でクラスターを形成しているようである。種々の根粒菌の生産するNodファクターの基本構造は共通で、キチン骨格、特にその還元末端残基を修飾する側鎖の化学構造のわずかな違いが、宿主マメ科植物に対する厳密な特異性を決定していることはよく知られている。そうした微妙なNodファクターの構造の違いを識別するために、受容体遺伝子はマメ科植物種間で多様化していなければならない。クラスターを構成する遺伝子ファミリーの内部で起こりうる相同組換えによって受容体遺伝子の多様化、言い換えれば宿主植物と根粒菌の間の厳密な特異性が獲得されたのかもしれない。 ミヤコグサがマメ科モデル植物として1992年に初めて提案されて以来、およそ10年近くに及ぶ分子遺伝学的な研究基盤の整備を経て、今ようやく植物−微生物共生の分子メカニズムに関する科学は大きなブレークスルーを迎えようとしている。向こう2年程度の間に、Nodファクター受容とそれに引き続くシグナル伝達系に関与する重要な植物遺伝子が次々と単離されるだろう。近い将来、これら遺伝子の産物の機能と相互関係が詳細な生化学的解析によって明らかになり、窒素固定共生をはじめとする植物−微生物共生の分子機構の全体像が解明されることを期待したい。 |
|||||||||||||||||||
|
||||||||||||||||||||
| 河内宏、梅原洋佐(農業生物資源研究所) | ||||||||||||||||||||
|
||||||||||||||||||||
| 2004年10月29日 | ||||||||||||||||||||
|
|
||||||||||||||||||||
|
||||||||||||||||||||

