|
Nod-factor を介したマメ科植物・根粒菌の相互作用
|
 |
|
 |
根粒菌(Rhizobium、Bradyrhizobium、Azorhizobium、Mesorhizobium、Sinorhizobium属)はマメ科植物の根に侵入し、根粒という新しい器官を形成して、共生窒素固定を行う。マメ科植物と根粒菌の関係は、一部の例外をのぞいて厳密な宿主特異性に支配されている。たとえば、S.melilotiはアルファルファに、B. japonicumはダイズに根粒を形成し、それらが交差することはない。こうした宿主特異性は、植物根から分泌されるフラボノイド等の化学物質を認識して根粒菌nod遺伝子群が活性化しリポキチンリゴ糖を合成・分泌する段階と、そのリポキチンリゴ糖を植物が認識・受容して根粒形成と感染のプロセスが開始する段階の、少なくとも2段階により決定されている(Fig.1)。宿主に根粒形成反応を引き起こすリポキチンオリゴ糖をNod-factor と呼ぶ。
|
|
|
| Fig. 1. 根粒菌と宿主植物の相互作用 |
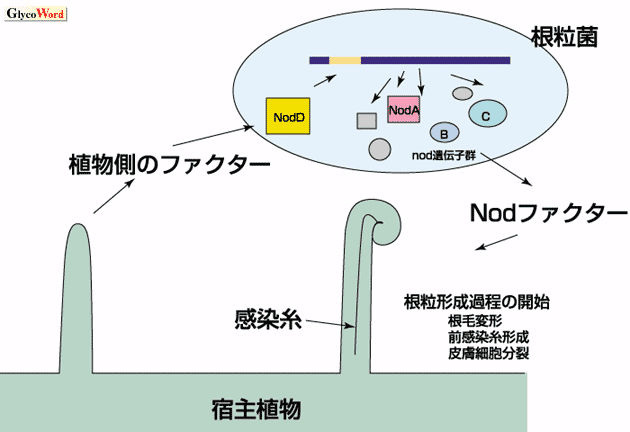 |
| 宿主特異性は少なくも2段階の植物微生物間の相互のシグナル交換により決定される。第一に植物から分泌されるフラボノイド等のシグナル分子に応答して、根粒菌側の根粒形成 (nod) 遺伝子が活性化し、Nodファクターの生合成と分泌が起こる。第二にNodファクターが宿主植物の根に根粒形成を誘導し、感染過程の引き金を引く。 |
|
|
|
Nod-factorは根粒形成と感染成立という2つの場面でシグナル物質として作用する。根粒菌から精製、または化学合成されたNod-factorは10-9〜10-12Mという低濃度で、宿主植物に根毛の変形、前感染糸形成や皮層細胞分裂を引き起こし、アルファルファや野生ダイズではほぼ正常な構造を持つ根粒形成を誘導することができる。
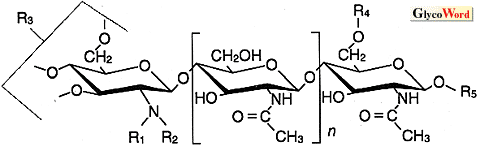
Nod-factorは、N-アセチル-D-グルコサミンがβ-1,4結合した3〜6 merを骨格とし、その非還元末端糖のC-2部位のNにアセチル基でなくアシル基(脂肪酸)が付加された基本構造をもつ(Fig. 2)。この脂肪酸は炭素数が16〜20、不飽和結合数が0〜4個と、様々な種類がある。Nod-factorの基本構造の合成はNod A、Nod B、Nod Cによって行われる。まずキチン合成酵素であるNodCにより基本骨格が合成され、NodBにより非還元末端の糖残基が脱N-アセチル化、最後にNodAにより、脂肪酸転移が行われて基本構造が完成する。nodA,B,Cは、転写調節因子であるnodDとともにすべての根粒菌に存在し、common(共通)nod遺伝子と呼ばれている。
|
|
|
|
| 根粒菌 |
宿主植物 |
R1 |
R2 |
R3 |
R4 |
R5 |
n |
|
| S.melioti |
M.sativa(アルファルファ) |
H |
C16:2 |
Ac(O-6) |
Sulfate |
H |
1,2,3 |
|
M.truncatula |
|
C16:3 |
H |
|
|
|
|
| R.leg.bv viciae |
Pisum(エンドウ) |
H |
C18:1 |
Ac(O-6) |
H |
H |
2,3 |
|
Vicia(カラスノエンドウ) |
|
C18:4 |
|
Ac(O-6) |
|
|
|
| R.leg.bv trifolii |
Trifolium(クローバ) |
H |
C18:2 |
Ac(O-6) |
H |
H |
2,3 |
|
|
|
C18:3 |
|
|
|
|
|
| M.loti |
Lotus(ミヤコグサ) |
Me |
C18:1 |
Cb |
AcMeFuc |
H |
3 |
|
|
|
C18:0 |
|
|
|
|
|
| R.fredii |
G.max(ダイズ) |
H |
C18:1 |
H |
MeFuc |
H |
1,2,3 |
|
G.soja(ツルマメ) |
|
|
|
Fuc |
|
|
|
| B.japonicum |
G.max |
H |
C18:1 |
Ac(O-6) |
MeFuc |
H |
3 |
|
|
|
C16:0 |
H |
|
|
|
|
|
|
C16:1 |
|
|
|
|
|
| B.elkanii |
G.max |
H |
C18:1 |
Ac(O-6) |
MeFuc |
H |
2,3 |
|
|
Me |
|
H |
Fuc |
Gro |
|
|
|
|
|
Cb |
|
|
|
|
| Me:メチル基, Ac:アセチル基, Fuc:フコシル基, Cb:カルバミル基, MeFuc:2-O-メチルフコシル基, Gro:グリセロール基 |
|
| Fig.2. Nodファクターの構造(reference2,改変) |
| NodファクターはNobBj-V(C18:1,MeFuc)の様に呼ばれている。BjはB.japonicum,Vはキチンオリゴマーの数を表す。()の中は、アシル基炭素鎖の長さと不飽和度、及び還元末端糖鎖の修飾基を示す。 |
|
|
|
|
|
Nod-factorの還元末端糖残基は、硫酸基やアセチル基、フコース、アラビノース等により、修飾を受けている。非還元末端にもN-アシル基とは別にカルバミル基やアセチル基が付加する場合もある。宿主特異性を支配するのは、基本骨格におけるアセチルグルコサミンの数及び、両末端糖の側鎖である。特に還元末端側の構造は重要である。これらの修飾に関与する根粒菌の遺伝子は宿主特異的nod遺伝子と呼ばれる。アルファルファを宿主とするS.melilotiは宿主特異的nod遺伝子nodH,Qによって還元末端に硫酸基のついたNod-factorを生産する。これらの遺伝子に変異を導入して硫酸基を付加できなくなった変異株はアルファルファに対する感染能を失うが、新たにベッチに対する感染能を獲得する。ダイズに感染するB.japonicumは宿主特異的nod遺伝子nodZによって還元末端にメチルフコースを付加し、メチルフコースのついたNod-factorだけがダイズに根粒を形成できる。非還元末端のアシル基の存在は生物活性に必須だが、その構造の違いは多くの場合、宿主特異性に決定的には影響しない。
植物側のNod-factor認識機構はほとんどわかっていない。根粒菌のnod遺伝子変異株や、化学合成した様々な構造のリポキチンオリゴ糖を用いた実験によって、植物によるNod-factor認識・受容は、特異性の異なる2つ以上のレセプターが関与するきわめて複雑なメカニズムによることが強く示唆されている。また、Nod-factorの受容の初期過程にレクチンが関与していることを裏付ける結果がある。Nod-factorの認識によって始動するシグナル伝達系についても、CaチャンネルやG-proteinの関与が示されているが、その基本的な性質はまったくわかっていない。しかし、植物側の認識機構に関する研究はまだ緒に付いたばかりである。生化学的な研究に加えて、Lotus japonicusやMedicago truncatulaといったモデル植物を用いた分子遺伝学的解析も開始されており、今後の進展が期待される。
|
|
|
|
|
|
梅原洋佐、河内宏(農業生物資源研究所・窒素固定研究室) |
|
|
|
|
|
| References |
(1) |
河内宏 (1997) 共生窒素固定と根粒形成のメカニズム 「分子レベルからみた植物の耐病性−植物と病原菌の相互作用に迫る」(山田哲治、島本功、渡辺雄一郎 監修) 細胞工学別冊 植物細胞工学シリーズ 8 pp.28-37 秀潤社、東京
|
|
(2) |
Spaink, HP : Regulation of plant morphogenesis by lipo-chitin oligosaccharides. Critical Reviews in Plant Sciences 15,559-582,1996
|
|
(3) |
Long, SR : Rhizobium symbiosis: Nod factors in perspective. Plant Cell 8,1885-1898,1996
|
|
(4) |
Denarie, J, Debelle, F, Prome, JC : Rhizobium lipo-chitooligosaccharide nodulation factors: Signaling molecules mediating recognition and morphogenesis. Annual Review of Biochemistry 65,503-535,1996
|
|
(5) |
Bladergroen, MR, Spaink, HP : Genes and signal molecules involved in the rhizobia-Leguminoseae symbiosis. Current Opinion in Plant Biology 1,353-359,1998
|
|
|
|
|
|
| 1999年 9月 15日 |
|
|
|
|
|
|



