

|
|
Evolution of Lactose and Milk Oligosaccharides |
|||||||||||||||||
 |
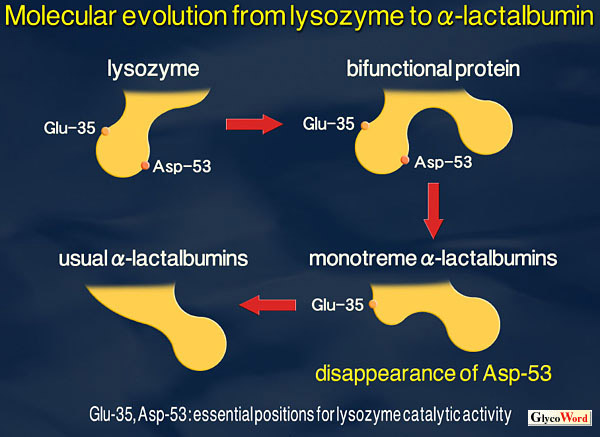
Mammalian milk or colostrum contains a few percent carbohydrates, of which disaccharide lactose (Gal( Although the amino acid sequence and tertiary structure of |
||||||||||||||||
 |
|||||||||||||||||
|
|||||||||||||||||
|
Mammalian milk or colostrum usually contains less than 20% of milk oligosaccharides, which consist of many saccharides with a lactose unit in the reducing end of the carbohydrate fraction, too, as well as lactose, while the milk of monotremes, marsupials, and bears, a eutherian species, contains more milk oligosaccharides than lactose. The heterogeneity of them has been found among the mammalian species. Human milk contains more than 100 oligosaccharides whose core units are classified into 12 groups, e.g., lacto-N-tetraose (LNT), lacto-N-neotetraose (LNnT), lacto-N-hexaose (LNH) and lacto-N-neohexaose (LNnH), etc. One hundred oligosaccharides are built by the attachment of Lewis a, b, x or
When the young of eutherians including human consume mother’s milk, the lactose is split into galactose and glucose by intestinal lactase, which is located in the membrane of the microvilli of the brush border of the small intestine, and the monosaccharides are transported into the enterocytes by a specific mechanism. The monosaccharides are used as energy sources in the young. The intestinal lactase is completely absent from the brush border of the villi of the small intestine of the suckling tammar wallaby. On the other hand, it is suggested that most milk oligosaccharides are not absorbed in human young, while they are regarded to be anti - infection factors to inhibit the attachment of pathogenic bacteria and virus in the colon. It is assumed that the milk oligosaccharides are absorbed in the small intestine of marsupial or monotreme young by pinocytosis or endocytosis and are then hydrolyzed by lysosomal glycosidases. It is thought that they are used as an energy source in the young and also act as anti-infection factors for them. Based on the above, the following hypothesis is proposed for the evolution of lactose and milk oligosaccharides. The proto-lacteal secretions of the primitive mammary glands of the common ancestor of mammals contained fat and protein but not lactose and milk oligosaccharides because of non-expression of |
|||||||||||||||||
| Tadasu Urashima (Obihiro University of Agriculture and Veterinary Medicine) | |||||||||||||||||
|
|||||||||||||||||
| Jul. 21, 2004 | |||||||||||||||||
|
|
|||||||||||||||||
|
|||||||||||||||||

