|
|
Mutants for Endosperm Starches in Rice |
 |
|||||||||||||
 |
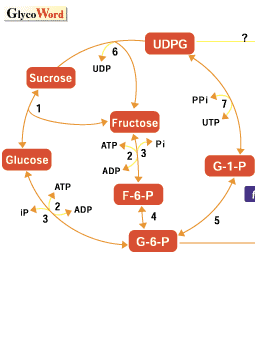
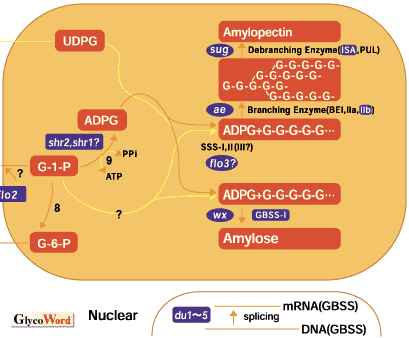
Every type of plant tissue forms an a D-glucose polymer, starch. It is produced as an end product of carbon fixation during photosynthesis and also accumulates in storage tissues as an important energy source in germination. Since Leloir et al. (1961) discovered the starch synthase reaction, the physiological mechanism of biosynthesis in starches in plants has been investigated intensively. However, some questions remain to be elucidated in the mechanism of genetic regulation in starch biosynthesis. In maize, various kinds of mutants for polysaccharides stored in maize grain are known, e.g. waxy, sugary, shrunken or opaque-2, etc. These mutants greatly contribute to the improvement of grain quality of maize, and thus expand the use of maize not only as the food for men and domestic animals but also for the important industrial materials in food chemistry. In addition to the importance in improving grain the quality of maize, they offer much valuable information in the study of gene action in the biological processes of metabolic regulation in higher plants. Excellent reviews and a book (1, 2, 3) treat these subjects in more detail. In rice, the mutants of endosperm properties were little known except for a waxy endosperm. However, other mutants have been found recently by mutation treatments. Although a wide variation of endosperm starches was observed in local rice cultivars collected from many countries, isogenic lines were helpful to examine the genetic and biochemical behaviors of a corresponding gene. As the artificial induction of mutation is one of the most effective methods to establish the isogenic lines by treating any cultivar, treatments using physical or chemical agents, for example, solar radiation such as x-rays or gamma-rays and ethyl methane sulfonate (EMS) have been employed toinduce mutations in plants. In addition, new biological techniques such as somaclonal mutation and transposon are available to introduce mutations in plants. We treated the fertilized egg cells at the mono cell stage after flowering in rice using N-methyl-N-nitrosourea (MNU) and obtained several thousands of rice mutants. The various kinds of rice mutants modifying endosperm starches quantitatively or qualitatively were found to be humerous as those found for maize. This method is characterized by the high frequency of mutation and a large size of mutated sector, making many mutations are easily detectable. In general, endosperm starches of cereals consist of two kinds of polysaccharides, amylose and amylopectin. Amylose is an essentially linear molecule consisting of alpha-1,4-glucosidic linkages, whereas amylopectin is highly branched with alpha-1,6-glucosidic bonds. The ratio of amylose to amylopectin has a major effect on the physical properties of starch. The proposed picture of starch today is a highly organized structure requiring sequential action of many distinct biosynthetic processes for its biogenesis. The enzymatic mechanism of starch synthesis can be considered to involve five processes: (1) synthesis of glucosyl-nucleotide substrate, (2) initiation of synthesis of unbranched maltodextrins, (3) elongation of alpha-1,4-glucan chains, (4) formation of alpha-1,6-glucan branches and (5) pre-amylopectin processing(trimming of alpha-1,6-branches), respectively. The metabolic functions of ADPG glucose pyrophosphorylase (AGPase), starch synthase (SS), starch branching enzymes (BE) and starch debranching enzymes (DBE), the possible key enzymes for starch biosynthesis, are production of glucan primer ADP glucose, elongation of alpha-1,4-glucan chains, formation of alpha-1,6-glucan branches and nonreducing ends as glucan acceptors, and trimming ofalpha-1,6-branches, respectively. Although amylose is synthesized by the enzymes responsible for the processes of (1) to (3), in which especially a granule-bound starch synthase (GBSS) involves in the elongation of alpha-1,4-glucan chains of amylose, the production of amylopectin is carried out with yhe cooperative action of different kinds of enzymes. In addition, plant tissues have multiple isoforms of respective enzymes. In rice there are at least three isoforms of starch synthase, one granule-bound starch synthase (GBSS) and two soluble starch synthase (SSSI and SSSII). Three isoforms of starch branching enzyme, BE I, BE IIa and BE IIb, are also recognized in rice. In addition, there are at least two kinds of isoforms of debranching enzyme, isoamylase (ISA) and pullulanase (PUL). The use of a variety of mutants in which the fine structure of amylopectin is differentially modified is indispensable to elucidate how each enzyme contributes to the construction of the amylopectin fine structure as shown in maize (Zea mays L.), pea (Pisum sativum L.) or a green alga, Chlamydomonas reinhardtii. In rice, various kinds of mutant for endosperm starch were generated by mutation treatments as follows. Wx gene encodes GBSS and thus waxy mutant caused by the lesion of Wx gene lacks or extremely decreases the amylose in endosperm starch. Dull mutation is characterized by low amylose content. Although five independent dull mutant loci have been so far identified (Yano et al., 1988), it is reported that one Du gene acts on the splicing of transcripted mRNA of GBSS gene (Wx gene)(Shimamoto et al., 2000). For the amylopectin, two kinds of mutations, amylose-extender (ae) and sugary 1 (sug 1), have so far been identified on the biochemical reaction of the biosynthesis. Genetic and biochemical analyses of these mutants strongly suggest that ae and sug 1 mutants encode a endosperm specific starch branching enzyme BE IIb and one of debranching-enzymes, isoamylase (ISA), respectively. Amylose-extender mutants produced most sparsely branched polyglucans, while sugary 1 mutants produced the most highly branched polyglucans called phytoglycogen. These structural alterations are reflected by the highest and lowest gelatinization temperature of the amylose-extender starch and the sugary 1 starch, respectively. The maize endosperm AGPase is composed of two immunologically distinct subunits, that is, large and small subunits. The maize mutants shrunken 2 (sh 2) and brittle 2 (bt 2) have kernels containing greatly reduced levels of ADP-glucose pyrophosphorelase compared to normal kernels. The biochemical and molecular genetic analysis of these mutants indicated that the large subunit of AGPase is missing in the endosperm of the sh 2 mutant and the small subunit in bt 2 and the sh 2 gene would be the structural gene for the large subunit, whereas the bt 2 gene would be the structural gene for the small subunit (Preiss et al., 1990). Rice endosperm mutants shrunken 1 (shr 1) and shrunken 2 (shr 2) also have kernels containing greatly reduced levels of ADP-glucose pyrophosphorelase compared to normal kernels. Details are under investigation. One of the most interesting mutants is a floury 2 (flo 2) mutant. The flo 2 mutants exhibited extremely reduced level of BEI in developing rice endosperm with decreasing levels of other starch-synthesizing enzymes including AGPase, GBSS, SSS I, BE IIb and PUL. Genetic and molecular analysis of the flo 2 mutants showed that the wild type Flo 2 gene co-regulates expression of the genes participating in starch synthesis in trans in a seed-specific manner (Kawasaki et al., 1996). Based on biochemical analysis of various mutants of rice endosperm, the sites of action of mutations are cited in Fig. 1. Only a few mutations of genes participating enzymes of starch biosynthesis have so far been identified and characterized genetically and biochemically, though we isolated more than one thousand mutants showing aberrant phenotype of endosperm in rice by MNU treatment. Considering the complicated pathway of starch biosynthesis in plants, many kinds of unknown mutants remain to be investigated. |
|||||||||||||
|
||||||||||||||
| Hikaru SATOH (Kyushu University, Faculty of Agriculture) | ||||||||||||||
|
||||||||||||||
| Sep. 15, 2000 | ||||||||||||||
|
|
||||||||||||||
|
||||||||||||||