|
|
Heparanase: A Heparan Sulfate-specific | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 | Heparan sulfate (HS) and heparan sulfate proteoglycans (HSPGs) are expressed by mammalian cells on cell surfaces and deposited in extracellular matrices including the basement membrane. Cell surface HSPGs play a role as receptors for adhesion molecules and growth factors; therefore they are implicated in cell adhesion, migration, differentiation and proliferation. HSPG is a real barrier of the basement membrane and functions as a filter against cationic molecules and macromolecules. HSPGs in the basement membrane also protect a major matrix component, type IV collagen from proteolytic attack. Cell surface HSPGs undergo continuous turnover in which an HS-specific endoglycosidase plays an important role to regulate intracellular metabolism of HSPGs. In blood borne metastasis, tumor cells disseminate from the primary site, intravasate, circulating in blood vasculature, and then extravasate at a distant organ site. Inflammatory cells also traverse a blood vessel wall and invade into tissues. These metastatic tumor and activated inflammatory cells penetrate the subendothelial basement membrane by enzymatic degradation of large components such as type IV collagen, laminin and HSPGs by a combination of proteases and endoglycosidases. HS-specific endoglucuronidase activity in a normal tissue was first reported by Höök et al. in 1975 (1). HS endoglycosidase activity was also found in fibroblasts, mast cells, and platelets. In 1980Ős a correlation between metastatic potential of tumor cells and their activity to degrade HS in the basement membrane were reported by many laboratories (2). An HS degrading glycosidase found in mammalian cells is an endoglucuronidase, which hydrolyses glucuronosyl glucosaminyl linkages in HS. This endoglycosidase is completely different from bacterial heparitinase, which is an eliminase; therefore we proposed to name the mammalian cell enzyme "heparanase"(3). Heparanase cleaves HS into characteristic large molecular-weight fragments, which need to be analyzed by gel permeation chromatography for detection and qantitation. Due to lack of a rapid assay suitable for a large number of samples for heparanase it took a long time to accomplish its purification. Almost 20 years have passed before the confirmation of a purified human heparanase by cDNA cloning and recombinant enzyme production, and five laboratories almost simultaneously published the purification and cDNA cloning of a human heparanase in the summer of 1999 (4-8). 1. Discovery of heparanase as an extracellular matrix degradation enzyme In late 1970's Nicolson and his colleagues tried to exploit the molecular mechanisms of organ-specific tumor metastasis and devoted their efforts to investigate tumor-endothelial cell interactions. They finally discovered that highly lung-metastatic mouse melanoma dells underlap vascular endothelial cells and degrade subendothelial basal lamina. The major degradation products released by melanoma cells from subendothelial matrix into culture media were HSPG carbohydrates chains, suggesting that melanoma possess endoglycosidase activity specific for HSPG (9). We further investigated this experimental model and found a good correlation between heparanase activity and lung colonization activity among mouse B16 melanoma sublines (10). The similar results were obtained with various metastatic tumor models such as mouse lymphoma (11) and rat mammary adenocarcinoma (2). An endoglycosidase found in mouse melanoma was identified as an endoglucuronidase similar to human platelet heparitinase (3). This enzyme was renamed ŇheparanaseÓ to distinguish itself from heparinase and heparitinase, eliminases produced by Flabovaterium heparinum (3). Incidentally, heparitin sulfate is now a dead term in glycosaminoglycan chemistry so that heparitinase is not a preferable name for the newly characterized enzyme specific for HS. The heparanase is highly active in acidic conditions (pH 4-6.5) suggesting that it is primarily a lysosomal enzyme. However, its activity was detected in cell membranes and in shedded vesicles. Tumor tissues often become hypoxic conditions by insufficient blood supply and undergo acidic conditions around pH 6. Thus, extracellularly released heparanases can easily function on the cell surface or in the extracellular matrix in such locally acidic conditions. An important feature of the mammalian cell heparanase is to recognize very unique domains in HS but not a simple repeat of four saccharides, IdUAGlcN(2-N-sulfate, 6-O-sulfate)-GlcUA-GlcN(2-N-sulfate, 6-O-sulfate) often shown in the biochemistry text books. 2. Purification and cDNA cloning of heparanase Heparanase activity is not found in stromal and epithelial cells in normal tissues. On contrast blood cells such as platelets and leukocytes possess heparanase activity. It is of interest that virally transformed cells and tumor cells show high levels of heparanase activity in association with malignancy. We used SV40-transformed human embryonic lung fibroblast WI38-VA13 for purification of heparanase (6). Heparanase has strong affinity to its inhibitor, heparin and is glycosylated with high-mannose-type polysaccharide chains. Thus, we used sequential column chromatography with heparin-sepharose and concanavalin A sepharose for purification of human heparanase to homogeneity (6). The purified heparanase showed a single peptide of 50 kDa detected by silver staining after SDS polyacrylamide gel electrophoresis. However, a fragment of 8 kDa derived from C-terminal domain of the proform heparanase also often coexists and could make a hetero dimer with 50-kDa active enzyme (8, 12). We analyzed N-terminal sequences of lysil-endoprotease digests of the purified heparanase, and searched for heparanase-matched cDNA sequences in the EST database. PCR primers for heparanase cDNA cloning were designed based on the sequences found in the EST database, and cloned a cDNA encoding human heparanase from a custom cDNA library of WI38-VA13 cells (6). The cloned cDNA has an ORF coding for 539 amino acid peptide with 6 potential N-glycosylation sites, which makes 65-kDa protein as a proform. The proform heparanase is converted by unidentified endopeptidases to 8 kDa peptide and 50 kDa catalytic domain with a lysine residue at the N-terminus. The expression of only 50-kDa peptide did not show heparanase activity, suggesting that a propeptide is necessary for formation of the active enzyme (4-8, 12). The gene of human heparanase is localized in 4q22 and composed of 14 exons and 13 introns (Table 1)(13). In WI38-VA13 cells alternative splicing produces two different size transcripts: 5 kb (HPSE1a) and 1.7 kb (HPSE1b) coding the same peptides (6). High levels of expression of the short transcript are detected in peripheral blood cells and placenta. All laboratories except for us employed a placenta cDNA library resulting in cloning of a short cDNA with 1.7 kb (4, 5, 7, 8). However, heparanases found in platelets and tumor cells were shown to be identical (5). | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
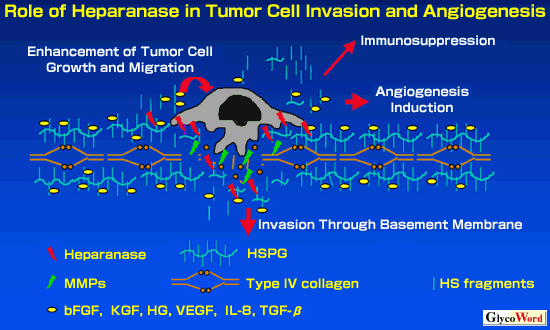
| 3. Heparanase as a new member of the Clan A glycohydrolase family All of the authors who reported their first purification and cDNA cloning of human heparanase described that there was no homologous proteins found in the databases; therefore heparanase is a unique protein (4-8). However, very recently, a previously undetected evolutionary connection between the heparanase and members of the TIM-barrel glycosyl hydrolase Clan A superfamily (GH-A) was found implying an unexpected structural and functional relationship (14). A unique domain of the catalytic site conserved among the plant glycohydrolases was detected in the human heparanase. Furthermore, two crucial amino acid residues conserved in GH-A were identified in the catalytic site of heparanase (14). Thus, heparanase was proposed to be a member of the new family 79, which is close to Family 10, 39 and 51 of GH-A (12). Heparanase is likely to contain an 8 (alpha/beta ) TIM-barrel fold, which is common to the GH-A. On the basis of sequence alignments with a number of glycosyl hydrolases from GH-A, two conserved acidic residues, Glu225 and Glu343 were identified as the likely proton donor and nucleophile residues, respectively, in a catalytic site of human heparanase (14). 4. Expression of heparanase in metastatic cancer and its prognostic value As described before, HSPG plays a role in regulation of various molecule transfer through the basement membrane, protection of matrix components such as type IV collagen from proteolytic attack, and storage of various cytokines and growth factors (2). HS degradation in the basement membrane causes release of angiogenic and growth factors leading to tumor cell migration, growth and angiogenesis, which are necessary for tumor cells to accomplish distant metastasis (Figure 1) (15). There have been ample evidence for a good correlation between metastatic potential and heparanase activity (2). It was also confirmed by the transduction study in which heparanase over-expession made tumor cells more metastatic in mice (4). In the clinical study a significant difference in heparanase gene expression level has been shown between different stages of cancer. We have observed a strong association of heparanase expression at both protein and mRNA levels with cancer progression of bladder carcinoma and urothelial cancer (16). Heparanase is over-expressed at the invasive edge of cancer tissue, and the extent of heparanase expression is correlated with poor prognosis (16). The patients with heparanase-negative urothelial cancer showed significantly higher disease-free survival than the patients with heparanase-positive cancer. It is of more interest that heparanase expression in urothelial cancer tissue was highly correlated with the number of angiogenic vessel in the tumor tiossues (16). In fact angiogenic endothelial cells express heparanase, and it has been also confirmed that activated fibroblast surrounding tumor cells, infiltrating macrophages as well as mast cells produce heparanase. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5. Inhibition of tumor metastasis by heparanase inhibitors A heparanase inhibitor reported at first was heparin (3). Mouse melanoma heparanase was active on HS of various origins but not active at all on any of chondroitin 4-sulfate, chondroitin 6-sulfate, dermatan sulfate, keratan sulfate, hyaluronic acid and heparin (3). Among these glycosaminoglycans tested only heparin inhibited heparanase. Heparin was already known as an inhibitor of tumor metastasis to the lung, and its activity was believed to be dependent on anticoagulation. We prepared anticoagulation activity-free heparin derivatives by chemical modification and yielded heparanase-specific inhibitors (17). Using these heparin derivatives we proved that heparanase inhibitors could suppress lung colonization of melanoma cells without influence on blood coagulation. There have been shown that lung colonization of metastatic tumor cells could be inhibited by heparanase inhibitors, including suramin (18), 6-O-sulfated carboxymethylchitin (19), and calcium spirulan (20). However, they are often also inhibitors of other biological activities such as growth factor binding and metalloproteinase activation (18-19). Sulfated phosphomannopentaose (PI88) was developed by Parish et al. as a heparanase inhibitor: it, however, does inhibit VEGF and bFGF binding to cell surface receptors. PI88 showed a good tolerability in Phase I study with healthy volunteers and subjected to Phase I study with cancer patients (12). 6. Additional comments All mammalian cells possess HSPGs on the cell surface and maintain them in a constant turnover by intracellular biosynthesis and degradation. They must constantly express heparanase at a certain level. In the contrary activated leukocytes express very high levels of heparanase. Thus a regulation of heparanase expression in activated leukocytes may be completely different from the one in normal stroma and epithelial cells. One of such mechanisms could be the expression of two different size transcripts, 1.7 kb and 5.0 kb, by alternative splicing. A shorter transcript is seen in peripheral blood cells and especially highly expressed in activated leukocytes. Inflammatory cells migrating from blood vessels to target tissue sites have to penetrate endothelial basement membranes and face to HSPG barriers, suggesting that heparanase activity is essential for inflammatory cells also. Therefore, a heparanase inhibitor could be useful not only for prevention of tumor metastasis and angiogenesis but also for suppression of inflammation. There is only one heparanase identified in mammalian cells so far. Hpa2 with a high homology to HPSE1 was very recently reported as a member of the novel heparanase family, but no heparanase activity has been detected with a Hpa2 recombinant protein (21). Thus, heparanase is a unique target enzyme for the development of new anti-metastatic and anti-inflammatory therapeutics. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Motowo Nakajima (Tsukuba Research Institute, Novartis Pharma K. K. ) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Jun. 30, 2001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||