 |
アトピー性皮膚炎は特定のアレルゲンによる慢性的な皮膚炎病態を示す疾患である。主な症状は搔痒、発疹、乾燥肌、患部の色素沈着である。特に激しい掻痒は集中力の低下を引き起こし、日常生活の多くの場面で支障をきたすため、QOLの低下の要因となる。アトピー性皮膚炎は小児で罹患率が高く、先進国においては20%の小児と10%の成人が罹患していると報告されている。現在、ステロイドなどを含む免疫抑制剤や抗体医薬などによる症状の緩和が主な治療法となっている (1)。
アトピー性皮膚炎はアレルゲンによる皮膚内の異常な免疫反応が原因である。特にヤケヒョウヒダニやコナヒョウヒダニなどのハウスダストマイト(House Dust Mite; HDM)が多くの患者において主たるアレルゲンであり、30 ~ 70%のアトピー性皮膚炎患者はHDMに対するパッチテストで陽性を示すと報告されている(2)。HDMは布団やソファー、絨毯のような高温多湿な環境に生息し、死骸やフンがアレルギー反応を誘発する。例えば、上述したダニのキチンやβ-グルカン、リポ多糖 (LPS)、セリンプロテアーゼなどが挙げられ、これらの分子は病原性関連分子パターン (Pathogen-Associated Molecular Patterns; PAMPs)として免疫細胞上のパターン認識受容体 (Pattern Recognition Receptors; PRRs)を刺激する。好中球、マクロファージ、樹状細胞などの自然免疫細胞はPRRのToll 様受容体 (Toll-Like Receptors; TLRs)を介してPAMPsを認識することで炎症誘発分子、サイトカインとケモカインを産生し、急性の皮膚炎を誘導する(図 1)(3)。この現象はアトピー性皮膚炎を増悪させる要因である。同じくPRRであるC型レクチン受容体 (C-type lectin receptors; CLR)はHDMや細菌を含めた病原体の糖鎖構造を認識する。CLRは細胞内シグナルを介して自然免疫細胞の活性を調整するが、一部のCLRはチロシンフォファターゼ-1/2 (SHP-1/2)を活性化し炎症反応を抑制することができる(図 2)。この現象は粘膜組織や皮膚において過剰な免疫応答を抑制し、組織の恒常性に重要な役割を果たす(4)。
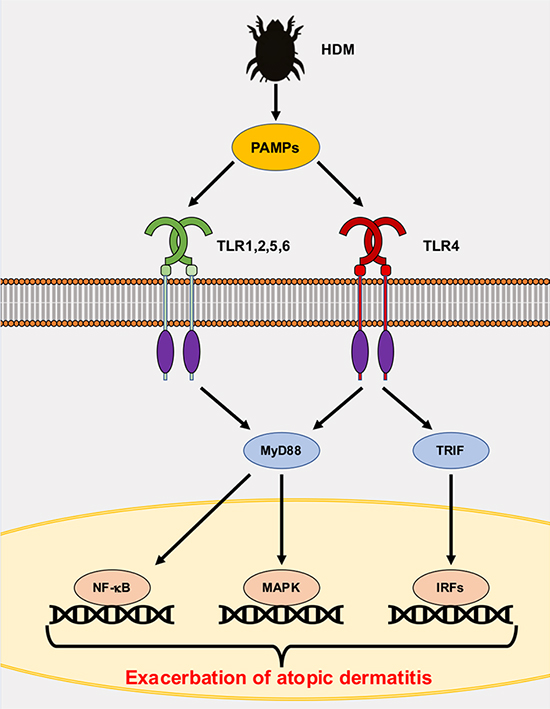
図 1. HDMによる炎症シグナルの誘導
HDM由来のPAMPsはTLRを刺激し、転写因子であるNF-κB, MAPK, IRFsを活性化する。これらの転写因子は炎症性サイトカインとケモカインの転写を促進し、炎症が誘導される。この反応はアトピー性皮膚炎を増悪させる。
|
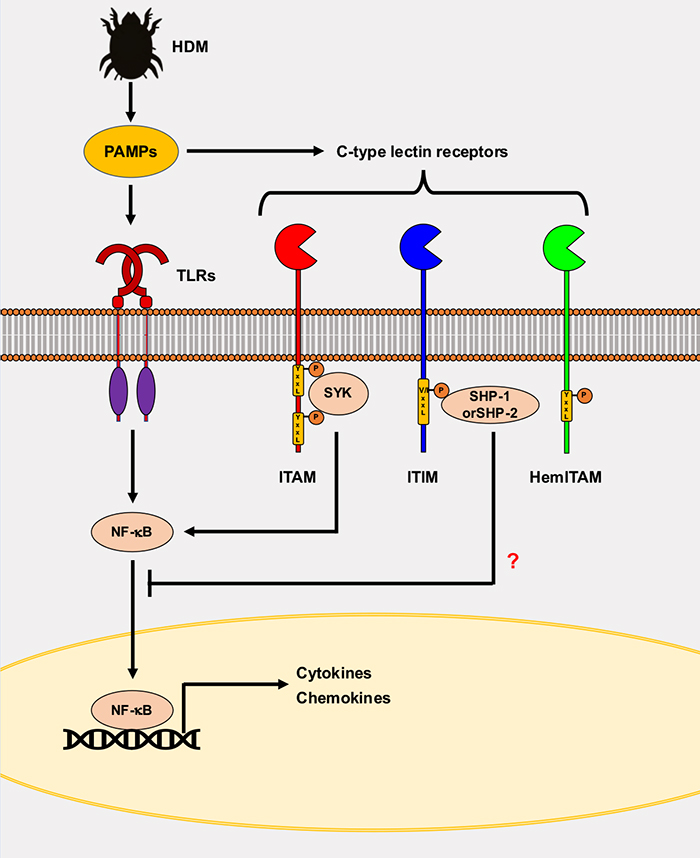
図 2. CLRの免疫学的役割
CLRの細胞内ドメイン上にあるITAM (immunoreceptor tyosine-based activating motif)は免疫活性を促進する。一方、ITIM (immunoreceptor tyosine-based inhibitory motif)は免疫活性を抑制する。一方HemITAMは活性化及び抑制のいずれかの機能を持つ。これらのアミノ酸配列は糖鎖認識ドメインにリガンドが結合するとチロシンがリン酸化し、下流シグナル分子を活性化させる。一部のCLRの下流シグナルはTLRsのようなPRRの下流シグナルに作用する。しかしながらチロシンフォスァターゼ (SHP-1/2)と会合するCLRがどのようにTLRの下流シグナルを抑制しているかは未だ不明である。
|
近年、CLRと様々な疾患との関連性が次々と明らかにされている(4)。例えば、CLRの一つであるDectin-2の遺伝子欠損マウスでは、野生型マウスと比較して、肝臓に接種した腫瘍細胞のクッパー細胞による貪食能が低下していることが報告された(5)。このことから、肝内のクッパー細胞はDectin-2を介して腫瘍細胞を貪食し肝転移を抑制することが示された。また、血小板に発現するClec2は、腫瘍細胞から産生されたポドプラニンと会合することで、血小板凝集を促進し、成長因子の放出や血管新生、免疫活性を抑制することが報告されている(6)。一方、自己免疫疾患やアレルギー反応との関連では、樹状細胞免疫受容体(DCIR)の多型が、関節リウマチの感受性と相関性があることが報告されている(7)。DCIRはチロシンフォスファターゼであるSHP-1を介して樹状細胞の過剰な活性化を抑制するが、DCIRの遺伝子欠損マウスでは樹状細胞の活性化が亢進し、自己免疫性炎症の自然発症することも示された(8)。また、鼻腔や気管支内の上皮細胞に発現するDectin-1は節足動物由来のトロポミオシンを認識すると、インターロイキン-33と呼ばれるサイトカイン産生を抑制し、アレルギー反応を抑制することが報告されている(9)。筆者らはマクロファージに発現する免疫抑制性CLRであるClec10aがHDMによって誘導されたアトピー性皮膚炎を抑制することを発見した(図 3)(10)。皮膚マクロファージはClec10aを介してHDM由来のムチン様分子を認識し、その結果、活性化したClec10aはHDMのPAMPsによって活性化したTLR4の下流シグナルを抑制し、炎症性サイトカインやケモカインの産生が低下させ、HDM誘発性皮膚炎を軽快させた。さらに、マウスClec10aのヒトのカウンターパートがヒトアシアロ糖タンパク受容体1 (hASGR1)であることを見出し、マクロファージに発現するhASGR1をノックダウンすると、HDMによるサイトカイン産生が増加したことから、hASGR1はマウスClec10aと類似した機能を持つことを明らかにした。
以上のように、近年CLR とアレルギーをはじめとした様々な疾患との関連が明らかにされてきている。CLRの分子標的薬開発を目的とした基礎研究と臨床開発研究はこれから更なる発展が期待される分野である。
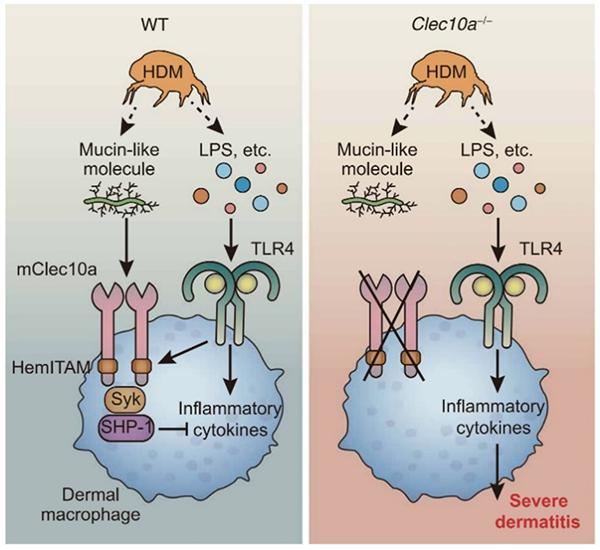
図 3. Clec10aの機能の概要図(Sci Immunol, 2019;4:eaax6908より引用)
マウスClec10aの細胞内ドメインにはHemITAMが存在し、免疫抑制を促すSHP-1と会合する。活性化したSHP-1はHDM由来の別のPAMPsによって活性化されたTLR4の下流シグナルを抑制する。その結果、マクロファージの免疫活性が減弱し、皮膚炎が抑制される。
|
重松 克伸 (筑波大学 医学医療系免疫制御学、ヒューマニクス学位プログラム)
渋谷 彰 (筑波大学 医学医療系免疫制御学、革新的創薬開発研究センター)
| References |
| (1) |
Ständer S: Atopic Dermatitis. N. Engl. J. Med. 384, 1136-1143, 2021 |
| (2) |
Darsow U, Laifaoui J, Kerschenlohr K, Wollenberg A, Przybilla B, Wüthrich B, Borelli S Jr, Giusti F, Seidenari S, Drzimalla K, Simon D, Disch R, Borelli S, Devillers AC, Oranje AP, De Raeve L, Hachem JP, Dangoisse C, Blondeel A, Song M, Breuer K, Wulf A, Werfel T, Roul S, Taieb A, Bolhaar S, Bruijnzeel-Koomen C, Brönnimann M, Braathen LR, Didierlaurent A, André C, Ring J: The prevalence of positive reactions in the atopy patch test with aeroallergens and food allergens in subjects with atopic eczema: a European multicenter study. Allergy 59, 1318-25, 2004 |
| (3) |
Jacquet A: The role of innate immunity activation in house dust mite allergy. Trends. Mol. Med. 17, 604-611, 2011 |
| (4) |
Brown GD, Willment JA, Whitehead L: C-type lectins in immunity and homeostasis. Nat. Rev. Immunol. 18, 374-389, 2018 |
| (5) |
Kimura Y, Inoue A, Hangai S, Saijo S, Negishi H, Nishio J, Yamasaki S, Iwakura Y, Yanai H, Taniguchi T: The innate immune receptor Dectin-2 mediates the phagocytosis of cancer cells by Kupffer cells for the suppression of liver metastasis. Proc. Natl. Acad. Sci. U S A 113, 14097-14102, 2016 |
| (6) |
Shirai T, Inoue O, Tamura S, Tsukiji N, Sasaki T, Endo H, Satoh K, Osada M, Sato-Uchida H, Fujii H, Ozaki Y, Suzuki-Inoue K: C-type lectin-like receptor 2 promotes hematogenous tumor metastasis and prothrombotic state in tumor-bearing mice. J. Thromb. Haemost. 15, 513-525, 2017 |
| (7) |
Guo J, Wu X, Too CL, Yin F, Lu X, He J, Li R, Liu X, Murad S, Padyukov L, Li Z: A replication study confirms the association of dendritic cell immunoreceptor (DCIR) polymorphisms with ACPA - negative RA in a large Asian cohort. PLoS One 7, e41228, 2012 |
| (8) |
Fujikado N, Saijo S, Yonezawa T, Shimamori K, Ishii A, Sugai S, Kotaki H, Sudo K, Nose M, Iwakura Y: Dcir deficiency causes development of autoimmune diseases in mice due to excess expansion of dendritic cells. Nat. Med. 14, 176-180, 2008 |
| (9) |
Gour N, Lajoie S, Smole U, White M, Hu D, Goddard P, Huntsman S, Eng C, Mak A, Oh S, Kim JH, Sharma A, Plante S, Salem IH, Resch Y, Xiao X, Yao N, Singh A, Vrtala S, Chakir J, Burchard EG, Lane AP, Wills-Karp M: Dysregulated invertebrate tropomyosin-dectin-1 interaction confers susceptibility to allergic diseases. Sci. Immunol. 3, eaam9841, 2018 |
| (10) |
Kanemaru K, Noguchi E, Tahara-Hanaoka S, Mizuno S, Tateno H, Denda-Nagai K, Irimura T, Matsuda H, Sugiyama F, Takahashi S, Shibuya K, Shibuya A, Fujisawa Y, Nakamura Y: Clec10a regulates mite-induced dermatitis. Sci. Immunol. 4, eaax6908, 2019 |
2023年 6月15日
|
|---|





