

|
|
Lectins Displaying RNase Activity (Leczymes) |
 |
||||||||||||||||
 |
Two sialic acid-binding lectins, isolated from eggs of Rana catesbeiana (cSBL) and R. japonica (jSBL), agglutinate a large variety of tumor cells, but do not agglutinate normal red blood cells and fibroblasts. This agglutinating activity is inhibited by sialoglycoproteins and gangliosides. SBLs are carbohydrate-free basic proteins consisting of 111 amino acid residues with 4 disulfide bonds. The sequence identity with cSBL is 80% for jSBL, 65% for RNase from liver of R. catesbeiana, 53% for onconase (anti-tumor protein from eggs of R. pipiens), 30% for angiogenin (potent inducer of angiogenesis) and 28% for RNase A, respectively. Because the residues His-12, Lys-41 and His-119, present in the active site of human pancreatic RNase are conserved in SBLs, and SBLs have pyrimidine-base specific RNase activity, SBLs belong structurally and functionally to the pancreatic RNase superfamily. Therefore, we call this class of SBL "leczymes". SBLs are cytotoxic proteins which inhibit the proliferation of tumors such as P388 and L1210 leukemia cells in vitro, and Sarcoma 180 and Ehrich ascites cells in vivo. Inhibition of tumor cell proliferation by cSBL is reduced by pretreatment of the cells with sialidase. cSBL binds to O-glycosidic oligosaccharide chains of sialoglycoprotein receptor(s) expressed on P388 cells, agglutinates the cells and is internalized into the cells via SBL receptor-mediated endocytosis. Internalized cSBL acts as RNase and induces RNA degradation leading to tumor cell death. cSBL-resistant clone RC150, an internalization-defective mutant of P388 cells, does not express the receptor involved in cSBL uptake, so that cSBL is not internalized into RC150 cells. Treatment of P388 cells with benzyl-GalNAc, which inhibits O-linked carbohydrate chain elongation, blocks cSBL-dependent cell death. Consequently, the initial steps in this process, binding of cSBL to the sialoglycoprotein receptor and its internalization, are important for the manifestation of anti-tumor effect of cSBL. cSBL can cause apoptotic morphological changes, such as nuclear condensation and disappearance of microvilli, to P388 cells (but not to RC150 cells). P388 cells are Fas antigen- and TNF receptor-negative. However, treatment with cSBL increases the expression of Fas antigen and TNF receptor on the cells. cSBL-treated P388 cells (neither RC150 cells nor benzyl-GalNAc-treated cells) display features of apoptosis, i.e., DNA fragmentation, binding of FITC-annexin V to the cell surface and activation of caspase-8 and -3 correlated with death factor (Fas ligand or TNF)-induced signaling pathway. Maximal activation of caspase-8 is detected at 3 h after treatment with cSBL. Addition of Ac-DEVD-CHO (inhibitor of caspase-3) or Z-IETD-fmk (inhibitor of caspase-8) [not Ac-YVAD-CHO (inhibitor of caspase-1)] inhibits the antiproliferative effect of cSBL. Pretreatment of cSBL with bovine submaxillary sialomucin inhibits cSBL-induced activation of caspase-3. The binding of cSBL to cSBL-sensitive P388 cells correlates with an increase rate of not only RNA but also DNA degradation and with the induction of apoptotic cell death. Caspase-3-dependent apoptotic cell death induced by cSBL raises an interesting question regarding the linkage between Fas antigen- or TNF receptor-dependent pathway and binding of cSBL to the sialoglycoprotein receptor. |
||||||||||||||||
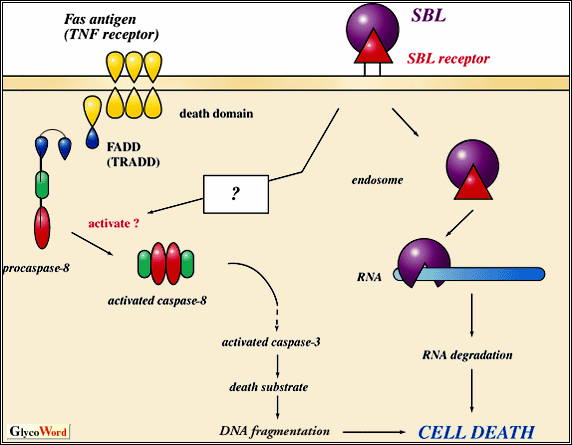 Fig : Mechanism of leczyme-induced cell death |
|||||||||||||||||
| Kazuo Nitta (Cancer Research Institute, Tohoku Pharmaceutical University) |
|||||||||||||||||
|
|||||||||||||||||
| Jun. 15, 1999 | |||||||||||||||||
|
|
|||||||||||||||||
|
|||||||||||||||||

