|
|
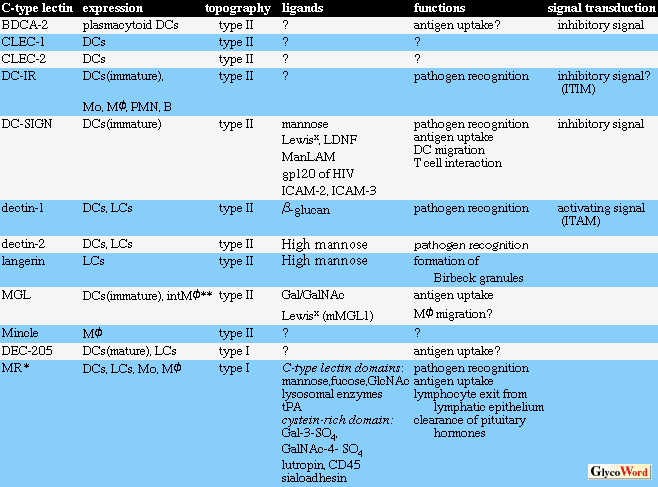
マクロファージ・樹状細胞に発現するC型レクチン |
||||||||||||||||||||||||||
| (First version published:2004年4月23日) | ||||||||||||||||||||||||||
 |
マクロファージ・樹状細胞は全身の末梢組織にくまなく存在しており、末梢から侵入する異物を殺傷、除去する自然免疫の担い手であるとともに、外来抗原を取り込んだのちに自身が移動してこれを二次リンパ器官へ運搬、Tリンパ球に抗原提示することにより獲得免疫を作動させる。近年、マクロファージ・樹状細胞に特異的に発現するC型レクチンが多数同定され(Table)、マクロファージ・樹状細胞の機能への関与について精力的に研究されている。C型レクチンは糖認識ドメインの構造のみならず、分子内に含まれる糖認識ドメインの個数、N末端の向き、細胞質領域のアミノ酸配列などに多様性を示す。またマクロファージ・樹状細胞には存在場所、分化段階、前駆細胞の異なる多くの亜集団が存在するが、C型レクチンにはlangerin、BDCA-2などのように特定の亜集団のよいマーカーとなる分子もある。そのリガンド分子は十分に解明されていない。なお、既知のC型レクチンの一部(L-selectinなど)もこの細胞集団に発現している。 |
|||||||||||||||||||||||||
|
||||||||||||||||||||||||||
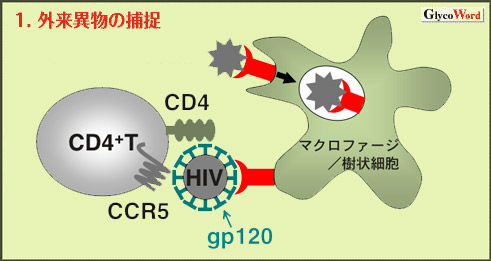
| 1.外来異物の捕捉 (図1) 近年同定されたC型レクチンの多くは外来異物の捕捉のための受容体として機能する。これらの分子は細胞質領域にチロシンモチーフ、ジロイシンモチーフなどのクラスリン被覆小胞へのシグナル配列、またリソソーム移行シグナルである酸性アミノ酸クラスターを有している。DC-SIGN (dendritic cell-specific ICAM-3 grabbing non-integrin)、マンノース受容体(MR)などいくつかのC型レクチンは種々の病原体を認識することができる。このC型レクチン依存的な取り込み機構はワクチン用の抗原を抗原提示細胞に能動的に送り込むために利用することができる。 HIVなどのウイルスが体内に侵入する際にはこのC型レクチンを侵入口として悪用する場合がある。ウイルスはまず末梢の粘膜組織に分布する樹状細胞上のC型レクチンに結合したのち、この細胞のもつ移動能を利用して二次リンパ器官へ移動・定着し、そこでTリンパ球に感染を広げるというモデルが提唱されている。HIV外被蛋白質gp120の結合蛋白質としてDC-SIGNが同定されており、樹状細胞からTリンパ球への感染がこの分子などを介して効率化されていることが指摘されている。一方、ランゲルハンス細胞に発現するlangerinはHIVの防御機構として機能することが最近報告された。 |
||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||
|
図1
|
||||||||||||||||||||||||||
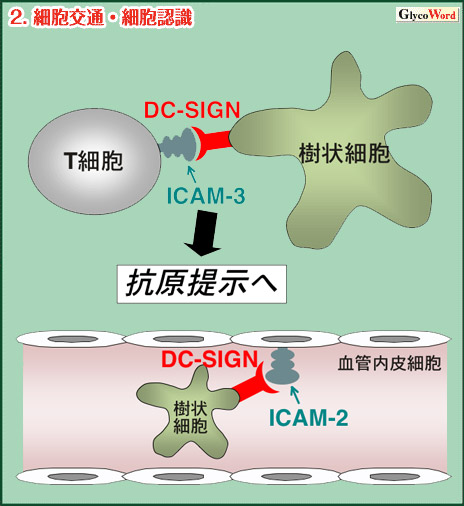
| 2.細胞交通・細胞認識 (図2) 樹状細胞前駆細胞が血管外遊走をする際には、樹状細胞側のDC-SIGNと血管内皮細胞側のICAM-2による初期接着が重要であると報告されている。 樹状細胞とTリンパ球との接着は抗原提示の成立に必須である。Tリンパ球側のICAM-3に対する認識分子としてDC-SIGNが同定された。DC-SIGNとICAM-3によるこの接着は細胞間相互作用の最も初期過程に関与している。 |
||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||
|
図2
|
||||||||||||||||||||||||||
| 3.シグナル伝達 C型レクチンの細胞質領域のアミノ酸配列は多様であるが、C型レクチンのいくつかはシグナル伝達に関与するモチーフを有し、実際に細胞の機能を調節する。 酵母細胞壁に由来するザイモザンはマクロファージ・樹状細胞の活性化物質として知られる。その構成物であるβグルカンの受容体としてdectin-1が同定された。dectin-1の細胞質領域にはITAM (immunoreceptor tyrosine-based activation motif) 配列が存在し、単独で、さらにToll-like受容体と共同して活性化シグナルを伝達することができる。dectin-1遺伝子欠損マウスの解析によりこの系が真菌に対する感染防御に寄与することが証明された。dectin-1は自然免疫系のシグナル伝達に関与する重要な認識分子である。 抑制シグナルを伝達する例も報告されている。plasmacytoid樹状細胞は抗ウイルス作用を示すIFN-αを大量に産生する。この亜集団に特異的に発現するBDCA-2は、インフルエンザウイルスの結合によって誘導されるIFN-αの産生に抑制的に作用するシグナル伝達分子であると考えられている。その糖鎖リガンドは未同定である。またDC-SIGNやMRはミコバクテリア由来のマンノース付加リポアラビノマンナン(ManLAM)と結合する。このManLAMはこれらC型レクチン経路を介してToll-like受容体による樹状細胞の成熟化を抑制すると考えられている。 |
||||||||||||||||||||||||||
| 東 伸昭(東京大学大学院薬学系研究科) | ||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
| 2007年5月22日 | ||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||
|
||||||||||||||||||||||||||