|
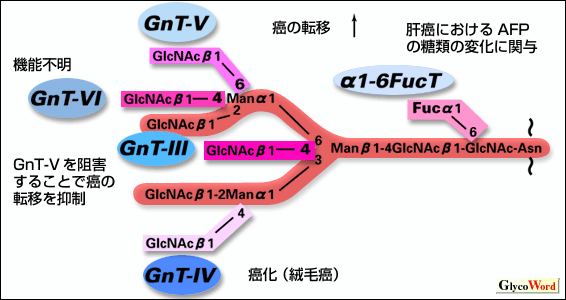
糖タンパク質の糖鎖構造は固有の糖転移酵素の作用により決定される。特に癌細胞では糖鎖の高分岐化が見られ(1)、転移能などの癌細胞の生物学的特性をモジュレートすることで近年注目されている(2,3)。高分岐化糖鎖構造の形成は、特定のKeyとなる糖転移酵素の活性化によって決定される。例えばN型糖鎖の高分岐化に関しては、下記に示すNアセチルグルコサミン転移酵素III, IV, V,VI (GnT-III, IV, V,VI)が分岐構造のコアとなる部分を決定するため、これらの酵素の活性化制御が全体の糖鎖構造の変化をきたす(4)。特にNアセチルグルコサミン転移酵素I, II (GnT-I, II)は基本となるコア構造の決定に(GnT-Iのノックアウトマウスは致死的)、シアル酸転移酵素やフコース転移酵素は糖鎖末端構造の決定に重要な意味をもつが、糖鎖の高分岐化の原因を最初に決めるのはGnT-III, IV, V,VIと言える。これらの酵素は組織特異的なプロモーターによって遺伝子レベルで制御され、さらにリン酸化などのpostotranslational regurationをうけて活性化される。面白いことに、これらの糖転移酵素は共通の基質を認識するにも関わらず、cDNAの構造にホモロジーがほとんど認められない。一般に、GnT-IVと GnT-Vは糖鎖の高分岐化に、GnT-IIIは高分岐化の抑制に作用すると考えられている。GnT-IVによって形成される絨毛性ゴナドトロピンの分岐構造は絨毛癌と胞状奇胎の鑑別に有用で、GnT-Vは癌の転移に関与すると言われている。特にGnT-Vに関しては、細胞増殖に伴う活性化、ets familyなどの転写因子による遺伝子制御、TGFβによるmRNAの安定化など複雑な制御をうける。糖鎖構造の決定には、in-vitroでの糖転移酵素活性のみでは説明できない部分がある。例えば、ある糖鎖構造にGnT-IIIが作用すると、糖鎖構造の立体障害により、もはやGnT-Vは作用しえなくなる。このことをin-vitroの系で利用した実験として、GnT-IIIの遺伝子導入によるメラノーマ細胞の肺転移抑制が報告されている。また近年、非還元末端のNアセチルグルコサミンにフコースを転移するα1-6フコース転移酵素 (α1-6FucT)がクローニングされたが(5)、この糖鎖構造は肝臓癌の腫瘍マーカーとして知られるα-fetoprotein(AFP)の癌性変化に関与すると同時に、末端の糖鎖構造の形成に対しても影響があると言われている。本酵素の過剰発現により、接着分子の機能が改変され、肝癌の実験的転移モデルにおいて転移抑制効果がみられた。 |
|
|
| References |
(1) |
Santer, UV, Gilbert, F, Glick, MC, Cancer Res. 44, 3730,1984 |
|
(2) |
Kobata, A, Eur. J. Biochem. 209, 483-501,1992 |
|
(3) |
Dennis,jw. Laferte, S. Waghorne, C. Breitman,ML. Kerbel,RS : beta 1-6 Branching of Asn-linked oligosaccaharides is directly associated with metastasis. Science 236, 582, 1987 |
|
(4) |
Taniguchi, N, Miyoshi, E, Ko, JH, Ikeda, Y, Ihara, Y : Implication of N-acetylglucosaminyltransferases III and V in cancer : Gene regulation and signaling mechanism. Biochem. Biophys Acta, 1999 in press. |
|
(5) |
Miyoshi, E, Noda, K, Yamaguchi, Y, Inoue, S, Ikeda, Y, Wang, W, Ko, JH, Uozumi, N, Li, W, Taniguchi, N : The alpha 1-6 fucosyltransferase gene and its biological significance. Biochem. Biophys Acta, 1999 in press. |
|
|
|
|
|