

|
|
スフィンゴ糖脂質合成阻害剤 | |||||||||||
 |
スフィンゴ糖脂質(GSL)の細胞機能に於ける本来の役割を探求するためには、細胞膜および細胞内に存在する内在性GSLを枯渇させることの出来る特異的GSL生合成阻害剤が有用である。GSLsの大部分は、セラミドとUDP-グルコ-スからUDP-glucose: N-acylsphingosine glucosyltransferaseにより生成されたグルコシルセラミド(GlcCer)を出発物質とし、これに種々の糖が付加されて生合成される。 セラミドの合成アナログであるD-threo-1-phenyl-2-decanoylamino-3-morpholino-1-propanol ( D-PDMP、図1) は、GlcCer生合成酵素の特異的な阻害剤であることがRadinと井ノ口により報告された(1)。 D-PDMPのGlcCer生合成酵素に対するIC50値は5μM, Ki値は0.7μMであり、阻害態度はセラミドに対して混合型競争阻害を示す。一方、ガラクトシルセラミド合成酵素やβ-グルコセレブロシダーゼ は阻害しない。 | ||||||||||
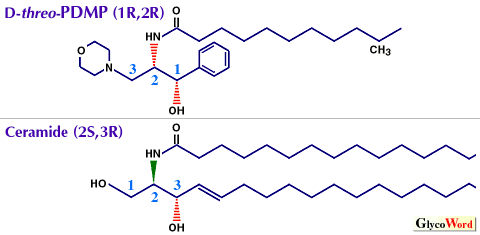 図1. PDMPとセラミドの構造比較 | |||||||||||
|
D-PDMPのGSL生合成阻害剤としての有用性は、今までに種々の培養細胞を用いて検討され、 濃度および処理時間に依存したGlcCer由来のGSLsの枯渇が可能であることが判明している。図1にPDMPとセラミドの構造を比較してある。天然型セラミドはD-erythro体であるのに対し、PDMPの4つの光学異性体の中でGlcCer生合成酵素阻害作用を有するのはD-threo体のみである。 D-PDMP処理した培養細胞では、細胞外マトリックス蛋白であるラミニンへの接着能の特異的減少、実験的肺転移の抑制、ブラジキニン刺激後のイノシトール3燐酸の生成亢進、IL2に依存したT細胞の増殖反応の抑制、Shope carcinomaに対する脱癌活性、シナプス形成抑制効果とシナプス形成必須因子としての機能性ガングリオシドGQ1bの同定等が報告されている(2)。これらの結果の解釈は、 D-PDMP処理細胞ではGSLs枯渇のみならず、グルコシルセラミドの前駆体であるセラミドやスフィンゴイド塩基の蓄積が起こることも考慮に入れて慎重になされるべきである。 PDMPのアナログとしては、アシル鎖の炭素数を変えた一連の化合物が合成され、炭素数14(PMMP)および16(PPMP)などは細胞内滞留時間がPDMPに比較して長いことから阻害効果の増強が期待される。また、フェニル部分をペンタデセニル基に置換したセラミドにより近い構造を有する類似体のGlcCer合成酵素に対する阻害活性について検討された結果、両者共にD-threo体に強力な阻害活性が確認されている。詳しくは最近のReview(2)をご参照戴きたい。 従来より、α-グルコシダーゼIの阻害剤として知られていたN-ブチルデオキシノジリマイシン(NB-DNJ)にGlcCer生合成酵素阻害活性があることが最近判明した。この発見が契機となって種々のN-アルキル化イミノ糖のGSL生合成に及ぼす効果が調べられた結果、N-ブチルデオキシガラクトノジリマイシン(NB-DGJ)はGlcCer生合成酵素の良好な阻害剤であり、細胞毒性も低いことが判明している。今後、NB-DNJおよびNB-DGJについてもそれらの阻害機構やゴーシェ病の治療薬としての検討も含めて研究が進展するものと期待される。 | |||||||||||
| 井ノ口 仁一 (北海道大学大学院薬学研究科) | |||||||||||
| |||||||||||
| 1999年 3月 15日 | |||||||||||
| |||||||||||

