

|
|
Callose Syndromes |
|||||||||||||||||
 |
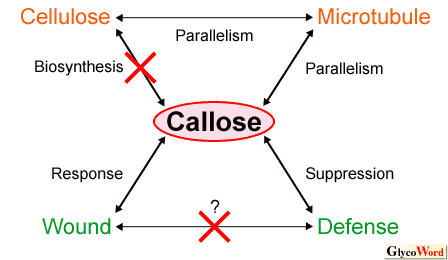
Cellulose-callose syndrome When we attempt to synthesize cellulose (1,4- Microtubule-callose syndrome Parallelism of microtubules to callose fibrils and loss of control over fibril orientation following microtubule disruption with propyzamide are characteristics of a syndrome that was observed in the glucan synthesis from UDP-glucose in the sheets of the plasma membrane (2). Callose synthase may bind to microtubules or microtubule-associated proteins directly or indirectly via other components of the enzyme complex. This syndrome is related to the fact that cellulose microfibrils are deposited parallel to cortical microtubules in plants. Wound response-callose syndrome Callose deposition induced by physiological stress, chemical and mechanical wounds, and pathogen infection of plant cells are characteristics of this syndrome, in which callose papillae barriers are believed to physically protect and impede pathogen attacks on plant cells. Although AtCSL5 (AtCalS12) is responsible for callose synthase related to wound response, its knockout mutant did not show a pathogen-induced callose formation but instead gained a phenotype of resistance to pathogens. Nishimura et al. (3) suggest that callose could be an induced defense response with negative feedback regulation by which callose synthesis suppresses the pathway of salicylic acid-dependent disease resistance. The salicylic- and pathogen-responsive genes are upregulated in the mutants, and these genes are hyperinduced after infection. In addition, Jacobs et al. (4) suggest that callose protects fungi during pathogenesis either by facilitating nutrient uptake or by preventing host-pathogen interactions. Of the many problems associated with callose, the major question remains its actual function in plants. |
||||||||||||||||
|
|||||||||||||||||
| Takahisa Hayashi (Research Institute for Sustainable Humanosphere, Kyoto University) | |||||||||||||||||
|
|||||||||||||||||
| Jun. 30, 2005 | |||||||||||||||||
|
|
|||||||||||||||||
|
|||||||||||||||||

