|
|
糖鎖が担う卵・精子間の種特異的な認識 |
|||||||||||||||||||||||||||||||||||
 |
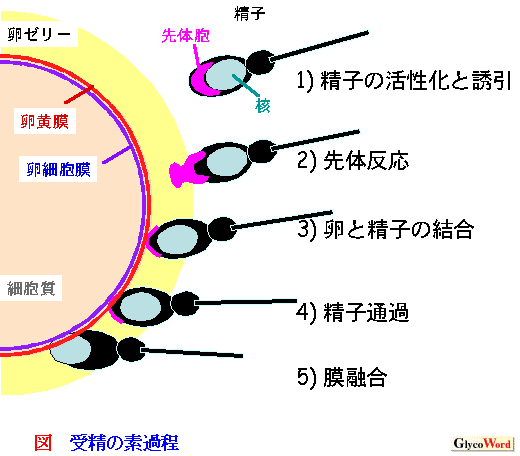
受精において精子と卵との間で精妙に相互認識することは、種の保存のために必須である。通常、卵は多くの糖蛋白質が含まれている卵外被に包まれているが、卵外被はバクテリアなどの外界からの攻撃から卵を守るために存在するばかりではなく、卵と精子が互いに同種の相手であることを認識しあう場という役割も持つ。精子は卵外被由来の物質に誘引され、卵外被と結合し、卵外被から受けたシグナルにより細胞内pHと細胞内Ca2+の上昇を起こし、精子の先端の先体胞のエキソサイトーシス(先体反応)を誘起し、卵外被を通過し、卵黄膜と結合、通過し、卵細胞膜へ到達する。このエキソサイトーシスによって露出した新しい精子細胞膜のみが卵細胞膜と融合することができる1)(図)。このような先体反応および、卵黄膜と精子の結合における細胞間相互認識には、糖鎖が重要な役割を持つことが知られている。 |
||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||
|
棘皮動物ウニの卵では、卵外被である卵ゼリー上に存在する巨大分子の硫酸フコースポリマー(FSP)が先体反応を誘起する。多種のウニにおいて種特異的なFSPの構造は決定されている2)。Strongylocentrotus franciscanus(米国産アカウニ)とS. purpuratus(米国産ムラサキウニ)では、α1→3結合したFSPの糖鎖に結合する硫酸基の位置に違いがある。前者のFSPではフコースの2位が硫酸化され、後者ではさらに4位が硫酸化されており、この違いをそれぞれの精子が認識し、種特異的に先体反応を誘起する3)。精子膜上にはFSPが結合する卵ゼリー受容体(suREJ1)が存在し、抗suREJ1抗体は先体反応を誘起することが可能である。SuREJ1のN末端にはほぼC型レクチンタイプのスーパーファミリーである糖認識ドメイン(CRD)が存在している。このようなCRDは、細胞認識、自然免疫、細胞シグナルに重要な役割を担っているドメインとして知られるため、suREJ1とFSPの結合にも寄与すると予想できる。Vacquierらは、6種のウニのsuREJ1-3のCRDを進化的に解析し、このドメインの進化速度が早く、それぞれの種が分化するように正の淘汰(Darwinian selection)を受けている遺伝子であることを示した4)。また同じ、棘皮動物のヒトデでは、卵外被の卵ゼリーに含まれる3種の分子が協調的に先体反応誘起に関わっている。キヒトデAsterias amurensisの場合、先体反応の誘起における主役の物質は巨大硫酸化糖蛋白質複合体の先体反応誘起物質(ARIS)であり、補助的にステロイドサポニンのCo-ARISと34アミノ酸残基のペプチドasterosapが働く。ARISはプロナーゼ消化により蛋白質を分解しても失活せず、過ヨウ素酸酸化により糖鎖構造をこわすと失活することから、糖鎖がその活性を担っている。その最小活性単位は直鎖型の5糖 →4)-β-Xylp(1→3)-α-Galp(1→3)-α-Fucp(4-SO3-)(1→3)-α-Fucp(4-SO3--)(1→4)-α-Fucp(1→ が10回くり返し構造をもつ糖鎖構造である5)。キヒトデと同じ科(Asteriidae)に属する近縁のヒトデの精子に対してキヒトデARISは同種と同様のレベルで先体反応を誘起するが、科が異なるヒトデの精子には誘起しない。キヒトデのARISのみならず同科の4種ヒトデ卵ゼリーでも同様な結果が得られた。同科のヒトデの卵ゼリーには、ARISと同じ硫酸基を含んだ5糖繰り返しの糖鎖構造が存在するが、科の異なるヒトデには存在しないことが示され、先体反応は科のレベルで特異的なARISの糖鎖の構造を認識していると考えられる。これらのことから、遺伝的な近縁種間では糖鎖およびその硫酸化が、生殖的隔離による種分化に貢献してきたのではないかと想像できる。
卵と精子の結合には、通常は糖鎖を修飾する酵素が、レクチンのように糖を認識結合する分子としてのみ作用している場合が知られている。原索動物カタユウレイボヤ,マボヤでは、卵外被である卵黄膜のフコース末端と精子表面のフコシダーゼ、Phallusia mammillataではGlcNAc末端とヘキソサミニダーゼとが結合することが受精に必須であることが知られている6-8)。 地球上に生息する数千万種の生物種にとって配偶子間の精緻な認識を可能にする唯一の機構が、糖鎖による認識機構であろう。 |
|||||||||||||||||||||||||||||||||||
| 松本 緑(慶應義塾大学理工学部生命情報学科) | |||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||
| 2006年3月31日 | |||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||