

|
|
N-結合型糖鎖の生合成と生物進化 |
||||||||
 |
マンノーストリミングの普遍性 |
|||||||
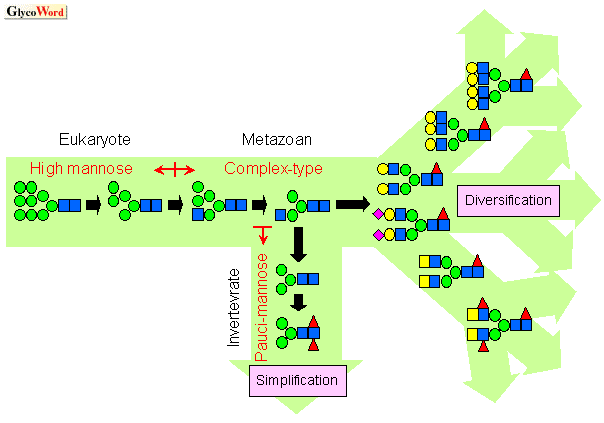 |
||||||||
|
図 |
||||||||
| 長束俊治(大阪大学大学院理学研究科) | ||||||||
|
||||||||
|
|
||||||||
| 2006年5月31日 | ||||||||
|
|
||||||||
|
||||||||
