

|
|
The Evolutionary History of Glycosyltransferase Genes |
|||||||||||
 |
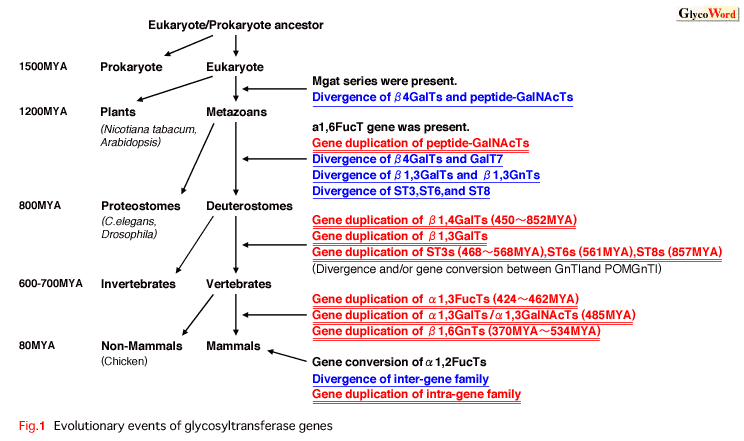
More than 160 human glycosyltransferases have been cloned to date. The number of cloned enzymes is increasing and may reach 200 within one year (Narimatsu, Glycoconj. J. 2004). Glycosyltransferases catalyze the transfer of sugar residues from nucleotide sugars (donor) to a growing carbohydrate chain (acceptor). Glycosyltransferases can be grouped into functional families based on their sequence similarities, which reflect their enzymatic characteristics: donor specificity, acceptor specificity, and linkage specificity between donor and acceptor. By the accumulation of glycosyltransferase gene data, it becomes evident that glycosyltransferases have strict specificity of linkage between donor and acceptor as well as donor specificity, whereas they show much lower specificity for acceptor. Here the general line of evolutionary history of glycosyltransferase gene families is discussed. The authors conducted molecular evolutionary analysis on some glycosyltransferase genes and mainly describe glycosyltransferase genes for N- or O-glycan synthesis (Kaneko, TIGG, 2000). The phylogenetic trees showed that the glycosyltransferase genes increased in number through gene duplication. We estimated the divergence time of each branch root and suggest that the glycosyltransferase genes increased in number through gene duplication and genome duplication (Fig. 1). The results show that gene duplication of the intra-gene family is concentrated around the early period of vertebrate lineage. Prior to gene duplication of the intra-gene family, gene duplication of the inter-gene family seems to have occurred. Briefly, in the ancestral genome, the ancestral glycosyltransferase may have multiplied by gene duplication and acquired variable activities via accumulation of point mutation, gene conversion, exon shuffling, and other phenomena. After the ancestral genome was equipped with a basic set of ancestral glycosyltransferase genes, genome duplication events may have occurred. |
||||||||||
 |
|||||||||||
|
Recent investigations have demonstrated that two genome-duplication
events occurred (tetraploidization); the first is close to the origin
of the vertebrates and the second is close to the origin of the gnathostomes
(Kasahara, Trend in genetics, 1997). We noticed that glycosyltransferase
genes show ?homologous cluster,? which was defined as more than two
chromosomal homologous segments. For example, the MHC gene clusters
were detected on chromosomes 1, 6, 9, and 19. Genes of the Hox clusters
were also mapped to chromosomes 2, 7, 12, and 17. These four regions
may be the remnant of the four homologous segments, called homologous
cluster that arose from the genome tetraploidization. |
|||||||||||
|
<Supplement> |
|||||||||||
| Mika Kaneko (Department of Experimental and Forensic Pathology, Yamagata University School of Medicine) Naruya Saitou (Division of Population Genetics, National Institute of Genetics) |
|||||||||||
|
|||||||||||
|
|
|||||||||||
| Mar. 14, 2005 | |||||||||||
|
|
|||||||||||
|
|||||||||||
