

|
|
概論:糖の化学進化と生物認識 |
||||||||||||||
 |
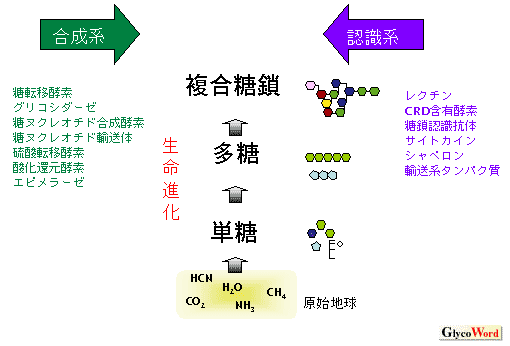
我々が糖鎖を研究するのにはおそらく三つの理由がある。第一に、糖鎖が生物にとって機能的に重要であるということ(機能的重要性)、第二に、核酸やタンパク質と比べてその合成や構造解析が格段に難しく、その解決が生命科学のパラダイムを切り開く可能性が高いこと(科学者にとって挑戦し甲斐のある課題)、そして第三に、糖鎖は生命の発祥とその進化と密接に結びついているということだ(生命起原・進化との関係)。もちろん、第三点については検証のしようがないが、もし糖鎖の起原が核酸・タンパク質と同等、あるいはそれらより古いとすると、糖転移酵素や糖ヌクレオチド合成に関わるタンパク質(糖鎖合成系)とこれらがつくる糖鎖を特異的に認識、あるいは相互識別するような糖鎖認識系(レクチン、サイトカイン、糖鎖認識抗体など)は生命進化の時を共有しながら互いに連携して進化してきたと考えられる(図1)。 | |||||||||||||
 |
||||||||||||||
|
||||||||||||||
|
本ジャンル「進化、比較生物学と糖鎖」では以下3つの項目に分け、糖鎖について「進化」、ないし「比較生物学」という切り口で、広範な生物現象を対象に解説を試みる。すなわち、「糖鎖の比較生物学」「糖鎖の生物進化」「レクチンの比較生化学と役割」というサブ項目の中でいくつかトピック性の高い用語を中心に、それぞれの分野で活躍される専門の先生方に解説して頂く。ここでは、先ずその大前提となる糖の化学進化について概説する。
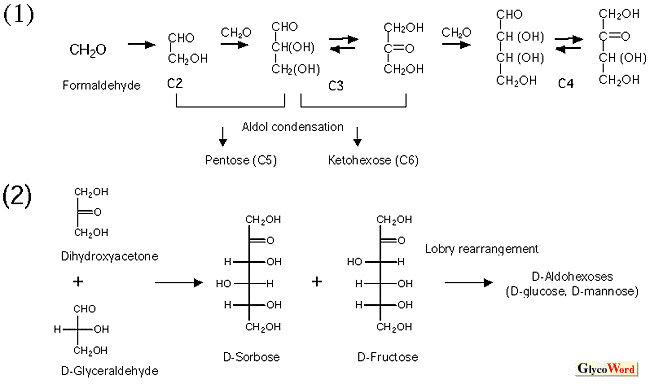
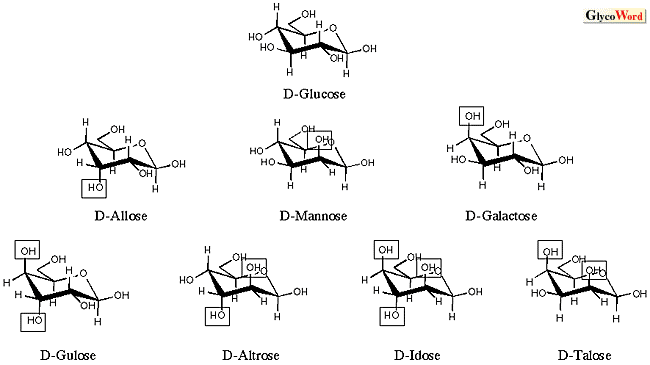
糖の起原・化学進化に関する論述(仮説)は、文献上筆者自身による論文(2)が唯一であるが、骨子を簡単に述べると以下のようになる。糖の非生物的合成(図2.ホルモース反応・アルドール縮合・Lobry de Bruyn転移)が可能であることから、生命誕生前に糖の化学進化があったと想定される。化学進化の時代を経て蓄積したフルクトース・グルコース・マンノースが生命起原時に利用可能な題材となった(生命起原三糖=First triplet)。生物進化の時代に入ると、原始生命は生命起原三糖を最大限活用し、様々な代謝系を発展させ、その結果数多くの後生糖を生み出した(生物進化によるブリコラージュ) 。この際、生物は最小限の労力で最大限の成果を得る戦略を選び(4-ケト中間体やアルドール縮合の頻用など)、かつすべて可能性のある糖(アルドヘキソース)を作り出すのではなく、ピラノース型が椅子型構造をとったとき十分安定な糖のみを選択して合成した。各構造の安定性はピラノース環構造を不安定化する1, 3-ジアキシャル相互作用の貢献によって判断できる(図3)。ガラクトースは高等動物において広範な認識現象に関わる中心的な糖であるが、図2に示した生命起原三糖の生成機構では生じ得ない(Lobry de Bruyn転移でガラクトースが生成するためには3-ケトースの存在が必要)。このため、ガラクトースは生物進化の後期になって登場した「後生糖」ではないかと類推される(ガラクトース・“late-comer”説)。因みに、フルクトース・グルコース・マンノースの相互変換は直鎖型の6-リン酸化体として起こるが、ガラクトースの生合成はNAD+を補酵素としてUDP-Glcの4-ケト化→4-エピメル化を経て行われる。前者は基本的に酵素無しにも起こる化学反応(Lobry de Bruyn転移に他ならない)だが、後者は補酵素無しには起こりえない複雑な反応で、明らかに前者とは異質の反応である。 |
||||||||||||||
 |
||||||||||||||
|
||||||||||||||
 |
||||||||||||||
|
||||||||||||||
| 平林 淳(独立行政法人 産業技術総合研究所 糖鎖工学研究センター ) | ||||||||||||||
|
||||||||||||||
| 2004年7月21日 | ||||||||||||||
|
|
||||||||||||||
|
||||||||||||||

