

|
|
Glycosylation of the AIDS Virus and an AIDS Vaccine |
||||||||||||||||
 |
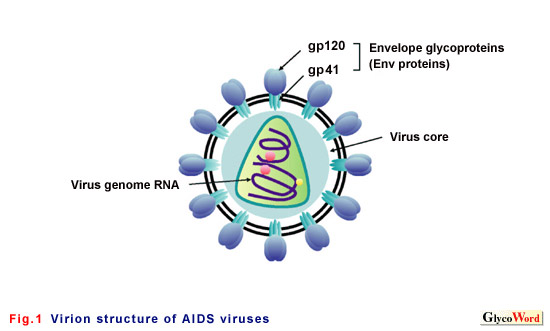
Envelope proteins of the AIDS virus (Env) that form viral spikes consist of gp120 and gp41 (Fig. 1). Gp120 comprises carbohydrates more than spike proteins of other viruses. Gp120 consists of a peptide with 500 amino acid residues and 22 to 26 N-linked glycans (Fig. 2). This structure of Env, wrapped in carbohydrate, is associated with the properties of the AIDS virus: evading host immune response in infected host and binding to target cells during infection. | |||||||||||||||
 |
||||||||||||||||
 |
||||||||||||||||
|
Evading host immune response
In virus entry, Env binds two receptors on target cells, CD4 and a chemokine receptor (CCR5 or CXCR4) (Fig. 3). Thus, Env is potentially a primary target of neutralizing antibodies (NAb). However, as described, most of the surface of the polypeptide part of Env is covered by abundant carbohydrate. Furthermore, the binding site for the chemokine receptor in gp120 critical for viral entry is hidden by loop structures formed in polypeptide part before binding to CD4. Therefore, effective NAb against AIDS virus is rarely induced. However, there have been reports that removal of parts of the carbohydrate improved induction of NAb (1) and that changes in N-glycosylation sites were associated with the emergence of neutralization escape mutants (2). |
||||||||||||||||
 |
||||||||||||||||
|
Pathogenicity of the AIDS virus
Twenty-three N-linked glycosylation sites reside in gp120 of a simian immunodeficiency virus (SIVmac239). We created a deglycosylation mutant . Cell tropism of AIDS virus From the in vitro study on viral properties, we found that Application of the deglycosylation mutant virus in AIDS vaccine development |
||||||||||||||||
| Chie Sugimoto
(CREST, Japan Science and Technology Agency) Kazuyasu Mori (AIDS Research Center, National Institute of Infectious Diseases Tsukuba Primate Research Center, National Institute of Biomedical Innovation) |
||||||||||||||||
|
||||||||||||||||
| Apr. 30, 2005 | ||||||||||||||||
|
|
||||||||||||||||
|
||||||||||||||||

